|
Nomi comuni:
Bangladesh |
bengali |
dahuk |
Bangladesh |
inglese |
goggle-eyed goby |
Cina |
cinese mandarino |
薄氏大弹涂鱼,
薄氏大彈塗魚 (Bo shi da tan tu yu -
grande saltafango di Bo) |
Filippine |
tagalog |
bia |
Finlandia |
finlandese |
nuoliliejuryömijä |
Germania |
tedesco |
Glotzauge |
Giappone |
giapponese |
mutsugoro |
Indonesia |
giavanese |
blodok, lunjat |
Italia |
italiano |
boleoftalmo di Boddart* |
Malaysia |
bahasa Malaysia |
ikan belacak binte biru |
Tailandia |
tailandese |
บู่ลาย, bulai |
Vietnam |
vietnamita |
cá bong sao, cá lác bo đa, cá thòi lòi |
* nome proposto
|
|
|
Sinonimi:
*In: Cuvier
& Valenciennes, 1837
Etimologia:
'Boleophthalmus' deriva dalla capacità che hanno questi pesci di elevare rapidamente gli occhi al di sopra livello delle orbite, come se "schizzassero" fuori dalle orbite: da 'bolê' (lanciato), e 'ophthalmôn' (occhio) (Cuvier & Valenciénnes, 1837)
la specie è dedicata a Pierre Boddaërt, che ha raccolto il materiale per la descrizione originale; 'boddaerti' è una correzione non giustificata (emendatio incorrecta) della versione originale del nome specifico (Murdy, 1989; Catalog of Eschmeyer)
|
Lunghezza massima registrata:
135 mm SL (Murdy,
1989)
220 mm TL (Kottelat et al., 1993)
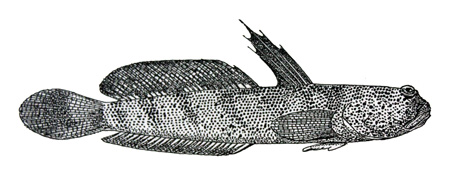
Colorazione in vivo (Murdy, 1989; Takita et al., 1999; Polgar & Crosa, 2009):
colorazione di fondo da marrone a verdastra sul dorso e sui fianchi, da bianco a grigia sul ventre, più scura dietro l'ano e sotto la testa; testa e tronco con molti piccoli punti bluastri ed iridescenti; chiazze da scure a nere sulla testa e sulla nuca; può essere presente una striscia scura che va dalla narice anteriore all'estremità dorsale dell'opercolo; 7-8
barre a sella diagonali da marrone scuro a nere sul dorso, che posteriormente si estendono di norma al di sotto della linea mediana laterale; D1 verdastra con numerosi punti celesti e con delle chiazze più chiare dello sfondo (diagnostiche) lungo il margine, dietro le prime 2-3 spine; D1 tipicamente gialla nei giovani; D2 verdastra, con serie di punti blu fra i raggi; pinna caudale da grigia a blu; pinna anale trasparente con una striscia scura inframarginale, fino a completamente scura; pinne pettorali con membrana da gialla ad arancione, con margine dorsale da marrone scuro a nero (diagnostico);
base muscolare delle pinne pettorali con numerosi punti sparsi da bianchi a bluastri; pinne pelviche parzialmente pigmentate in posizione dorsale, biancastre e pigmentate solo prossimalmente in posizione ventrale
Colorazione in liquido conservante (Murdy,
1989; oss. pers.: Bangladesh, Peninsular Malaysia, India, Indonesia):
colorazione di fondo da marrone scuro a grigio scuro dorsalmente e lateralmente, biancastra fino a grigia ventralmente, con labbra e mento da grigio scuri a bluastri; bande a sella dorsali e chiazze scure sulla testa talvolta conservate; D1 e D2 marrone bluastro con punti biancastri; pinna caudale marrone fino a grigio scura; pinna anale biancastra con una striscia scura inframarginale fino a completamente scura; pinne pettorali grigiastre con margine dorsale pigmentato; pinne pelviche da biancastre a grigie
Diagnosi (Murdy,
1989; Takita et al., 1999; Polgar & Crosa, 2009):
elementi totali di D2 24-26; elementi totali della pinna anale 24-26; conta longitudinale delle scaglie 61-79; scaglie predorsali 25-35; lunghezza della pinna caudale 17.9-23.3%SL; lunghezza della testa 25.0-30.4%SL; lunghezza della base di D2 40.2-46.4%SL; primo elemento di D2 di norma non segmentato e non ramificato; denti della mandibola incisi (bifidi).
Chiazze più chiare dello sfondo sul margine dorsale della membrana di D1, dietro le prime 2-3 spine; margine dorsale delle pinne pettorali nerastro.
Dimorfismo sessuale specie-specifico: spine di D1 più allungate nelle femmine adulte (Polgar & Crosa, 2009; Swennen et al., 1995).
Il genere è caratterizzato dall'epidermide della testa, della nuca e del dorso particolarmente ispessita e ricoperta di papille dermiche, e da una cartilagine rettangolare che ricopre il cinto pelvico (vedi disegno)
Dieta:
questa specie è un erbivoro che si ciba sul fondo raschiando la superficie del fango con movimenti laterali della testa (diatomee bentiche,
alghe verdi: Khoo, 1966, citato in Clayton, 1993; vedi filmato). Dove è simpatrica con la congenerica B. pectinirostris (per es. Sud-Est Asia: Polgar & Crosa, 2009), gli studi non hanno preso in considerazione alcuna possibile differenza fra queste specie, a lungo confuse (B. dussumieri sensu Takita et al., 1999 e Polgar & Khaironizam, 2008 = B. pectinirostris sensu Polgar & Crosa, 2009). Lo stesso accade in India occidentale, dove B. boddarti è simpatrica con B. dussumieri (Murdy, 1989). Ad ogni modo, tutte queste specie hanno comportamenti alimentari pressochè identici ed è probabile che la loro dieta sia simile
Riproduzione:
i maschi saltano per attrarre le femmine all'interno delle tane (oss. pers.: Malaysia peninsulare); i dettagli del ciclo vitale non sono disponibili in letteratura, ma sono probabilmente simili a quelli delle specie congeneriche (vedi Comportamento riproduttivo e B. pectinirostris)
|
|
|
Note ecologiche:
localmente molto abbondante lungo il margine marino e nella zona a pneumatofore delle foreste a mangrovie (Polgar & Crosa, 2009); gli adulti scavano le tane sulle rive fangose di insenature tidali, foci di fiumi e su piane fangose, di norma non troppo lontano dalla prima fila di alberi (Takita
et al., 1999).
Durante la bassa marea, B. boddarti mostra una partizione ecologica fra individui di maggiori dimensioni, distribuiti in zone non coperte da vegetazione e vicino al bordo dell'acqua, ed individui più piccoli sul suolo della foresta e nella zona a pneumatofore; un pattern simile è stato osservato in habitat diversi dello stesso ecosistema in Periophthalmus gracilis ed Oxuderces dentatus, mentre un pattern apparentemente inverso è stato osservato in P. variabilis (Polgar & Bartolino, 2010).
Lo stesso pattern è stato osservato in Boleophthalmus poti
(= Boleophthalmus sp.; Polgar et al., 2010;
Polgar et al., 2013) in Papua New Guinea, and in
B. birdsongi in Australia
(Takita et al., 2011). Il ciclo vitale è stato studiato da Ravi e Rajagopal (2007)
a sinistra: Kuala Selangor, Malaysia peninsulare:
margine marino della foresta, rive fangose di un'insenatura tidale; tipico habitat degli adulti di
B. boddarti (foto: G. Polgar, 1996)
a destra: Tg. Piai, Malaysia peninsulare: piana tidale superiore con alcuni alberi di Rhizophora mucronata; qui la specie è molto abbondante (foto : G. Polgar, 2006)
|
|
|
|
|
Distribuzione:
regione indo-pacifica occidentale, dalle coste occidentali dell'India (Mumbai, Maharashtra) verso est fino al Sabah (Borneo settentrionale) e Vietnam meridionale; località tipo: Oceano Indiano (Murdy, 1989; Bucholtz & Meilvang, 2005)
|
Note:
questa specie è allevata estensivamente e largamente consumata in Tailandia (vedi sopra) (Clayton, 1993).
Chew & Ip (1992), Chew et al. (1998) and Ip et al. (2004c) hanno trovato una considerevole resistenza di questa specie alla presenza di cianuro (prodotto da diversi organismi, quali cianobatteri, funghi, piante, artropodi ed alghe: Ghosh et al., 2006) e solfuri (abbondanti in sedimenti anossici). Entrambi questi composti tossici inibiscono la respirazione aerobica cellulare attraverso l'inibizione della citocromo ossidasi. Come prima, rapida risposta adattativa, B. boddarti riduce la permeabilità dei tessuti alla NH3 e incrementa la produzione di NH3, aumentando così il pH dei fluidi corporei e convertendo gli acidi tossici nelle forme ionizzate, che hanno una minore permeabilità attraverso le membrane cellulari (Chew et al., 1998). In seguito, risposte enzimatiche specifiche nel fegato e nei muscoli mediano la detossificazione attraverso la produzione di composti non tossici: tiocianati, solfati, tiosolfati e solfiti in condizioni normossiche. In condizioni ipossiche, B. boddarti adotta meccanismi O2-indipendenti, detossificando i solfuri a zolfo solfani e al contempo sopprimendo l'attività della solfuro ossidasi, liberando così la riserva limitata di O2 (anche a causa dell'affinità relativamente elevata per l'ossigeno dell'emoglobina dei saltafango) per il metabolismo energetico aerobico (Ip et al., 2004c).
Tali risultati suggeriscono la presenza di adattamento evolutivo alle difese chimiche anti-predatorie adottate da microalghe e cianobatteri, di cui la specie si nutre, e alle elevate concentrazioni di solfuri nei fanghi anossici dove questi pesci scavano le tane.
E' interessante notare che solo esemplari di B. boddarti >8 g di peso corporeo (circa 80 mm SL: Khaironizam & Norma-Rashid, 2002) sono in grado di tollerare l'esposizione ai solfuri (Ip et al., 2004c). Ciò suggerisce che i giovani (SL< 80 mm) non siano in grado di fare uso delle tane anossiche durante l'alta marea, il che è in accordo con la presenza dei giovani a livelli topografici intertidali superiori rispetto agli adulti (Polgar & Crosa, 2009; Polgar & Bartolino, 2010), dove il substrato viene sommerso per tempi inferiori.
|
Fotografie di Boleophthalmus
boddarti:
|
|
A: una femmina di B. boddarti: si notino le spine allungate di D1 (foto: G.
Polgar, Kuala Lumpur, Malaysia, 2006); B: un giovane esemplare con D1 gialla (foto: G. Polgar, Kuala Lumpur, Malaysia, 2006);
C: un primo piano di una femmina (foto: G. Polgar, Kukup jetty, Malaysia peninsulare, 2006); D: un altro primo piano di una femmina (foto: G. Polgar, Kukup
jetty, Malaysia peninsulare, 2006); E: un incontro agonistico lungo i confini del territorio (foto: G. Polgar, Kukup jetty, Malaysia peninsulare,
2006); F: un individuo mentre "setaccia" in una piccola pozza la miscela di fango ed alghe raccolta dalla superficie del fango: si notino le increspature sulla superficie dell'acqua create dal rapido movimento della mandibola (foto: G. Polgar, porto di Kukup town, Johor, Malaysia peninsulare, 2006); G: un altro primo piano (foto: Akinori Kamiya
"Yamaneko", Can Gio, Viet Nam, 2004; © umisuzume 2006)*; H: B. boddarti predato dal serpente colubride Cerberus rynchops (foto: K. Khor, Kuala
Gula, Malaysia, 2005); I: un primo piano di due maschi che combattono (foto: K. Khor, Kuala Gula, Malaysia, 2005) - *
con il permesso dell'autore
|
Disegni di Boleophthalmus boddarti:
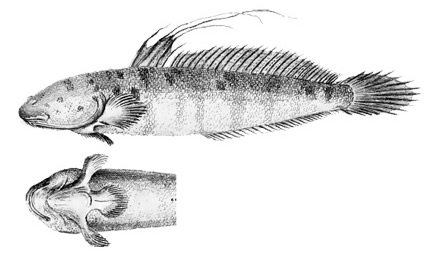
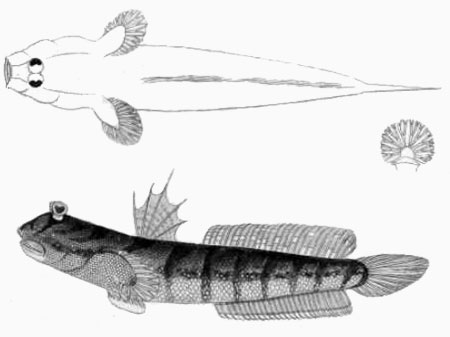
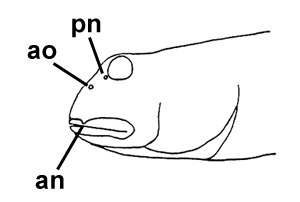
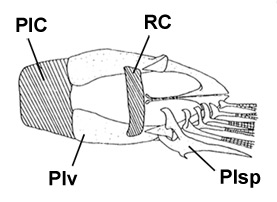
A: Boleophthalmus boddarti (Pallas) (Koumans, 1953); B: Gobius striatus Bloch & Schneider (Bloch & Schneider, 1801); C: B. boddaerti (Pallas) (Bleeker, 1983)*; D: Gobius boddarti Pallas (modificato da Pallas, 1770); E: Boleophthalmus boddaerti (Pallas) (Day, 1876; si noti che Apocryptes punctatus Day, 1867 non riporta illustrazioni, ma la descrizione corrisponde a B. boddarti); F: Gobius plinianus (Hamilton, 1822); G: pori cefalici sensoriali e nasali di Boleophthalmus spp.: an = narice anteriore; ao = poro del canale oculoscapulare anteriore; pn = narice posteriore (modificato da Murdy, 1989)*; H: vista ventrale del cinto pelvico di B. boddarti (gli elementi della pinna pelvica sono stati rimossi dal lato sinistro: PIC = cartilagine pelvica intercleitrale; Plv = pelvi; RC = cartilagine rettangolare; Plsp = spina della pinna pelvica (modificato da Murdy, 1989)* - * con il permesso dell'autore
|
Video di Boleophthalmus boddarti:

clicca sull'immagine per scaricare (tasto destro del mouse--> Salva con nome) o riprodurre
(tasto sinistro del mouse) il videoclip.
Un adulto di B. boddarti mentre si ciba: il pesce raschia il biofilm formato dalle diatomee bentiche premendo la mandibola sulla superficie del fango e muovendo la testa da un lato all'altro. Il raschiamento è facilitato dalle file di denti bifidi sulla mandibola, disposte su un piano quasi orizzontale (clip di Yuko Ikebe, Samut Prakarn, nei pressi della foce del fiume Chao Phraya, Tailandia centrale, 2001)
|