|
Nomi comuni:
Cina, Taiwan |
cinese mandarino |
大弹涂鱼, 大彈塗魚 (da tan tu yu - saltafango gigante) |
Finlandia |
finlandese |
loistoliejuryömijä |
Giappone |
giapponese |
mutsugorô |
Italia |
italiano |
boleoftalmo asiatico gigante* |
Malaysia |
Bahasa Malaysia |
ikan belacak binte biru |
Vietnam |
vietnamita |
cá lác |
* nome proposto
|
|
|
Sinonimi:
Etimologia:
'Boleophthalmus' deriva dalla capacità che hanno questi pesci di elevare rapidamente gli occhi al di sopra livello delle orbite, come se "schizzassero" fuori dalle orbite: da 'bolê' (lanciato), e 'ophthalmôn' (occhio) (Cuvier & Valenciénnes, 1837)
'pectinirostris' è una forma composta latina,
da 'pecten' (pettine, rastrello) e 'rostrum' (becco, muso), che probabilmente si riferisce ai denti incisi della mandibola
|
Lunghezza massima registrata:
155 mm SL (Murdy, 1989)
Colorazione in vivo (Murdy, 1989; Polgar & Crosa, 2009):
colorazione di fondo grigio verdastra; testa ricoperta da numerosi piccoli punti celesti, che su fianchi e dorso sono più grandi e diradati;
7 chiazze nere sui fianchi e all'attaccatura della pinna caudale, al di sopra della linea mediana laterale; negli adulti la cute al di sotto delle orbite (coppe dermiche) è celeste chiaro (popolazioni malesi). D1 grigio scura-verdastra con numerosi piccoli punti celesti (nei giovani non si osserva un cambiamento di colore come in B. boddarti). D2 grigio verdastra con colonne di 3-8 punti celesti fra i raggi;
pinna caudale da grigio verdastra a grigio scura con numerosi punti o chiazze allungate celesti sulla membrana interradiale; pinna anale con pigmentazione da arancione a grigio scuro nella porzione prossimale, distalmente da ialina a gigiastra; base
muscolare delle pinne pettorali con numerosi punti celesti; pinne pettorali grigiastre; pinne pelviche da grigiastre a grigio scuro
Colorazione in liquido conservante (Murdy, 1989;
oss. pers.: Malaysia peninsulare, Cina meridionale, Giappone meridionale):
colorazione di fondo da biancastra a marrone scuro; pinne dorsali e caudale grigio-bluastre; tutte le altre pinne marrone chiaro; i punti si conservano di solito solo
sulle pinne; in alcuni esemplari, 1-4 bande scure diagonali sono visibili sul dorso sulla parte posteriore del corpo; in alcuni esemplari si osservano grandi
chiazze scure sulla testa
Diagnosi (Murdy, 1989;
Polgar & Crosa, 2009):
elementi totali di D2 23-26; conta longitudinale delle scaglie 84-123; scaglie predorsali 26-48; lunghezza della pinna caudale 18.3-22.2%SL;
lunghezza della testa 24.3-28.0%SL; lunghezza della base di D2 41.5-46.1%SL;
primi elementi di D2 di norma segmentati e ramificati; denti della mandibola incisi (bifidi).
Malayan populations: negli adulti, la cute sotto le orbite (coppe dermiche) è di colore celeste; dimorfismo sessuale:
spine di D1 più allungate nelle femmine; nei maschi sono presenti chiazze di colore più scuro dello sfondo lungo il margine della
membrana di D1, dietro le prime 2-3 spine.
Il genere è caratterizzato dall'epidermide della testa, della nuca e del dorso particolarmente ispessita e ricoperta di papille dermiche,
e da una cartilagine rettangolare che ricopre il cinto pelvico (vedi disegno)
Dieta:
si ciba sul fondo ed è erbivoro, nutrendosi di associazioni di microalghe (Yang, 1996; Yang et al., 2003), che raschia dalla superficie del fango con movimenti laterali della testa, setacciando poi la miscela di acqua e fango presso uno specchio d'acqua (vedi foto J). Anche la flora batterica
dell'intestino di questi pesci è stata oggetto di studio (Morii &
Kasama, 1989; 1990).
Tutte le specie di questo genere mostrano comportamenti alimentari pressochè identici (e.g. filmato di B. boddarti) ed è probabile che anche la loro dieta sia assai
simile
|
|
Riproduzione:
i maschi saltano per attrarre le femmine all'interno delle tane.
Nelle femmine adulte delle popolazioni giapponesi la seconda spina di D1 allungata (MudskipperWorld, by Yuko Ikebe). Analogamente, nelle popolazioni malesi le femmine presentano spine di D1 più allungate che nei maschi (Polgar & Crosa, 2009). Ciò si osserva anche in esemplari malesi e tailandesi di B. boddarti (Polgar & Crosa, 2009; Swennen et al., 1995), e in esemplari di B. caeruleomaculatus della Nuova Guinea (Polgar et al., 2010).
B. pectinirostris è stata riprodotta artificialmente tramite ovulazione indotta ed allevamento delle larve
(Chen & Ting, 1986;
Chung et al., 1991;
Hong et al., 1988;
Hong & Wang, 1989;
Zhang et al., 1987;
Zhang et al., 1989;
Zhang & Zhang, 1988).
L'età e la crescita sono state studiate attraverso l'analisi dei radiali delle pinne pettorali
(Washio et al., 1991;
Nanami & Takegaki, 2005)
a sinistra: una femmina
a destra: un maschio
(foto modificate da MudskipperWorld, © Yuko Ikebe)
|
|
|
|
Note ecologiche:
localmente molto abbondante su piane tidali fangose aperte.
Le popolazioni malesi (Polgar & Crosa, 2009; Polgar & Bartolino, 2010) non sono chiaramente distribuite secondo una partizione ecologica fra differenti classi di taglia lungo la zona intertidale, come quella osservata in differenti habitat dello stesso ecosistema in B. boddarti, Oxuderces dentatus e Periophthalmus gracilis (Polgar & Bartolino, 2010); tuttavia, gli individui di maggiori dimensioni sono esclusivamente distribuiti sulla piana tidale inferiore, non coperta da vegetazione, mentre quelli più piccoli sembrano avere una distribuzione più irregolare, sia sulla piana tidale esposta che lungo corsi d'acqua effimeri all'interno del mangrovieto e nella zona a pneumatofore (Polgar & Bartolino, 2010).
Durante l'inverno, i pesci che vivono nel Giappone meridionale vanno in ibernazione sul fondo delle tane
(Takegaki et al., 2006)
a sinistra: tipico habitat di B. pectinirostris: si possono vedere le aperture delle tane (Giappone,
prefettura di Saga, Mare di Ariake)
a destra: calco in resina di una tana di un adulto: si noti la forma ad "Y" corrispondente alle due aperture; l'intero cunicolo
è profondo circa 1.5 m
(foto: modificate da Hiro Masa
Matsumoto, 1999)
|
|
|
|
|
Distribuzione:
Taiwan, China e Giappone meridionale;
località tipo: Canton, China (Murdy, 1989).
Di recente è stata rinvenuta in Asia sudorientale (Malaysia peninsulare) e in Indonesia
(Polgar & Crosa, 2009),
dove si trova in simpatria con B. boddarti.
Le diverse popolazioni malesi, cinesi e giapponesi mostrano colorazioni e forme lievemente differenti (vedi sotto: Note)
a sinistra: un esempio di "architettura-saltafango"
a destra: decorazione lignea ispirata al "mutsugorô"
(foto: Hiro Masa Matsumoto: prefettura di Saga, Giappone, 1999)
|
|
|
|
Note:
questa specie è largamente pescata ed allevata in Giappone, Taiwan e Cina meridionale, dove è consideata una delicatezza culinaria
(Clayton, 1993). In Giappone questo pesce è molto popolare,
e viene catturato con un metodo tradizionale chiamato
mutsukake,
tramandato da padre in figlio. Gli attrezzi più importanti sono una canna molto lunga, un amo a forma di arpione
ed una sorta di slitta da fango chiamata oshi-ita. Un'altra tecnica consiste nell'uso di trappole che imitano il cunicolo del pesce,
ricavate dal bambù, e che vengono conficcate nel fango. Secondo Polgar e Crosa
(2009), esemplari malesi di questa specie sono stati
erroneamente identificati come B. dussumieri da
Takita et al., 1999 e Polgar & Khaironizam, 2008
Takegaki (2008) ha sollecitato azioni urgenti per salvare le popolazioni del Giappone meridionale di questa specie,
minacciate dall'intensa distruzione del suo habitat in questa regione (Stanley, 2011).
Analisi filogenetiche molecolari suggeriscono che questa specie includa due specie criptiche con diversa distribuzione geografica: una nel Mar Cinese Meridionale (dal Golfo di Tonchino allo Stretto di Taiwan) e nel Mar Cinese Orientale fino allo stretto di Corea (B. pectinirostris s.s.), e l'altra nello Stretto di Malacca (la cui distribuzione potrebbe estendersi verso nordest fino a Taiwan: oss. pers.)
(Chen et al., 2014).
a sinistra: amo (foto di Hiro Masa Matsumoto, 1999)
nel centro: trappole tradizionali giapponesi (foto di Hiro Masa Matsumoto, 1999)
a destra: un pescatore su di una oshiita che va a pesca di mutsugoro
(© Ariake lawsuit, 2007)
|
|
|
|
Fotografie di Boleophthalmus pectinirostris:
|
A: B. pectinirostris in acquario (foto: © fishing-forum,
prefettura di Kumamoto, Giappone, 2003); B: una femmina da Taiwan (foto: Hans Ho, mercato di Taipei, Taiwan, 2006); C: un maschio da Taiwan (foto: Naomi
Delventhal, mercato di Taipei, Taiwan, 2006); D: un esemplare cinese: si noti la differente colorazione (foto: anonimo, Meizhou, China, 2006); E: uno scontro fra
due maschi territoriali; F: una femmina; G: maschi riproduttivi che saltano per richiamare l'attenzione
delle femmine (foto E-G: © BlueNativeFactory,
Mare di Ariake, Giappone, 2006)*; H: disputa territoriale fra due maschi; una femmina è fuori fuoco in primo piano (foto: G. Polgar, porto di Kukup town, Johor,
Malaysia peninsulare, 2006); I: un'altra foto di un maschio che salta, proprio nel momento in cui si spinge con la coda
(foto: Yuko Ikebe, Mare di Ariake, Giappone,
2003)*; J: un individuo mentre "setaccia" in una piccola pozza la miscela di fango ed alghe raccolta dalla superficie del fango: si notino le increspature
sulla superficie dell'acqua create dal rapido movimento della mandibola (foto: G. Polgar, porto di Kukup town, Johor, Malaysia peninsulare, 2006); K: un esemplare
comprato al mercato (foto: Ray Cui, Guangzhou, Cina, 2006)*; L: una femmina di B. pectinirostris e un maschio riproduttivo di
Periophthalmus chrysospilos - si noti l'entrata della tana di
quest'ultima specie nell'angolo in basso a destra (foto: G. Polgar, Tanjung Piai, Malaysia Peninsulare, Johor, 2007) - *con il permesso dell'autore
|
Disegni di Boleophthalmus pectinirostris:
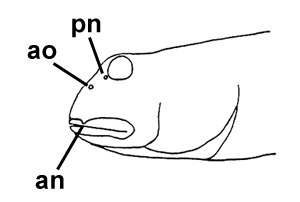
A: pori cefalici sensoriali e nasali di Boleophthalmus
spp.: an = narice anteriore; ao = poro del canale oculoscapulare anteriore; pn = narice posteriore
(modificato da
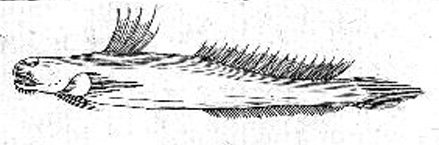
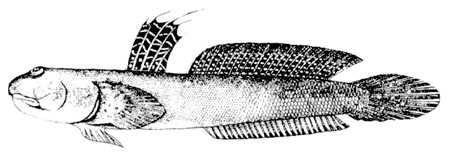
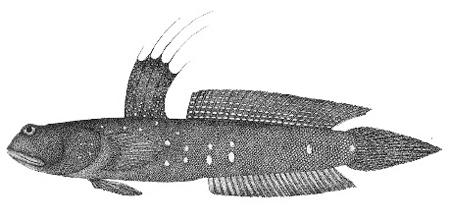
Murdy, 1989)*; B: Gobius pectinirostris Linnaeus, 1758: il primo disegno scientifico di un saltafango (Linnaeus, 1759: dettaglio della fig. 3, p. 260) Göttinger Digitalisierungs - Zentrum, Goettingen State and University Library*; visitato: 08/2007); C: un esemplare coreano (Kim, 1997);
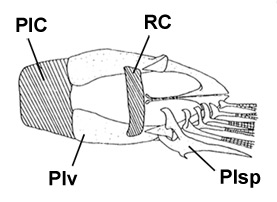
D: vista ventrale del cinto pelvico di B. boddarti (gli elementi della pinna pelvica sono stati rimossi dal lato sinistro:
PIC = cartilagine pelvica intercleitrale;
Plv = pelvi; RC = cartilagine rettangolare; Plsp = spina della pinna pelvica (modificato da Murdy, 1989)*; E: Boleophthalmus pectinirostris (Linnaeus) (Bleeker, 1983)*; F: Boleophthalmus chinensis (Osbeck) (Herre, 1927) - * con il permesso dell'autore
|