Nomi comuni :
Cina |
cinese mandarino |
杜氏大弹涂鱼 () |
India, Gujarat |
gujarati |
લેવતી (Levti) |
Iran, Hormozgan |
farsi (persiano) |
eshlambo, neeshlambo, gel khorok |
Iraq, Oman |
arabo |
kelb-el-daw, abou-shlembo, abu-shelamboo |
Italia |
italiano |
boleoftalmo di Dussumier* |
Pakistan |
urdu |
gullo |
* nome proposto
|
|
|
Sinonimi:
Boleophthalmus dussumieri |
Valenciénnes, 1837* |
(sin. senior, combinazione originale) |
Boleophthalmus dussumierei |
Valenciénnes, 1837* |
(emendatio incorrecta) |
Boleophthalmus dentatus |
Valenciénnes, 1837* |
(sinonimo junior) |
Boleophthalmus chamiri |
Holly, 1929 |
(sinonimo junior) |
Pseudapocryptes dentatus |
vari autori |
(combinazione non valida) |
*In: Cuvier & Valenciénnes, 1837
Etimologia:
'Boleophthalmus' deriva dalla capacità che hanno questi pesci di elevare rapidamente gli occhi al di sopra livello delle orbite, come se "schizzassero" fuori dalle orbite: da 'bolê' (lanciato), e 'ophthalmôn' (occhio) (Cuvier & Valenciénnes, 1837)
la specie è intitolata al viaggiatore e mercante francese Jean Jacques Dussumier, che raccolse il materiale tipo (Murdy, 1989)
|
Lunghezza massima registrata:
187 mm TL (Randall, 1995)
289 mm TL (la maggiore registrata in un gobide oxudercino: Abdoli et al., 2009)
Colorazione in vivo (Murdy, 1989;
oss. pers.: Iran, Hormozgan):
colorazione di fondo da grigio-bluastra a marrone chiaro su dorso e fianchi, con numerosi punti marrone scuro sul lato dorsale della testa, opercoli, guance, porzioni muscolari delle pinne pettorali, e sul tronco; colorazione ventrale più chiara;
barre scure verticali a sella irregolari su dorso, testa e nuca possono essere visibili, ma mai al di sotto della linea mediana laterale;
D1 grigio chiara con numerosi punti scuri sulla membrana interradiale;
D2 grigia con colonne di punti azzurro chiaro sulla membrana interradiale (più grandi in posizione basale), e pochi punti neri in posizione anteriore e basale; uno sottile margine scuro può essere presente, con un bordo arancione;
pinna caudale con raggi da grigio scuri a nerastri, e membrana interradiale azzurra;
pinna anale ialina; pinne pettorali da marrone chiaro a celeste chiaro con serie di punti più scuri lungo i raggi; pinne pelviche marrone chiaro, più scure in posizione prossimale
Colorazione in liquido conservante (Murdy, 1989;
oss. pers.: Iran, Iraq, Kuwait):
colorazione di fondo da marrone chiaro a grigio-bluastra fino a violacea; i punti più scuri su corpo e pinne possono essere visibili; barre a sella sul dorso assenti; colorazione di fondo di D1 e D2 da grigiastra a marrone scuro; D2 con punti bianchi e margine scuro spesso visibili; pinna caudale da grigiastra a marrone scuro; pinna anale e pelviche ialine; pinne pettorali da marrone chiaro a ialine
Diagnosi (Murdy, 1989):
elementi totali di D2 24-28;
lunghezza della pinna caudale 21.9-25.5%SL;
elementi totali della pinna anale 24-27;
conta longitudinale delle scaglie 103-185;
conta delle scaglie predorsali 48-56;
lunghezza della testa 23.6-28.7%SL;
lunghezza della base di D2 42.2-44.7%SL; primo elemento di D2 di norma non segmentato e non ramificato; denti bifidi sulla mandibola. I tre canini superiori su entrambi i lati della sinfisi sono allungati, e sporgono in maniera cospicua in alcuni esemplari conservati; dimorfismo sessuale:
i maschi hanno una D1 più alta e canini più lunghi (Berg, 1949, citato da Brian Coad: visitato il 10/2009).
Il genere è caratterizzato dall'epidermide della testa, della nuca e del dorso particolarmente ispessita e ricoperta di papille dermiche, e da una cartilagine rettangolare che ricopre il cinto pelvico (vedi disegno)
Dieta:
Pankow & Huq (1979) e Sarker et al. (1980) hanno descritto il contenuto stomacale di Pseudapocryptes
dentatus da Shatt al-Arab, Iraq; Salih & Al-Jaffery (1980)
hanno studiato alcuni caratteri morfologici in relazione al comportamento alimentare di B. dentatus (Iraq). Questa specie si ciba principalmente di diatomee epibentiche, ma anche di cianobatteri, alghe filamentose, insetti, crostacei, nematodi ed uova di teleosteo. Il comportamento di "grazing" (raschiamento del substrato), con il tipico movimento oscillatorio della testa, è molto simile a quello delle altre specie congeneriche (vedi filmato di B. boddarti).
Riproduzione:
Hoda & Akhtar (1985) e Hoda
(1986) hanno studiato la maturazione sessuale e la fecondità di B. dentatus
e B. dussumieri (Pakistan, costa di Karachi) considerandole specie simpatriche. Il comportamento di corteggiamento è stato descritto da Clayton
& Vaughan (1988): i maschi saltano per corteggiare a distanza le femmine, in maniera non dissimile alle altre specie congeneriche (oss. pers.). A Karachi (Pakistan), i giovani e i cosiddetti "young-of-the-year" si trovano da settembre a dicembre;
la maturazione sessuale inizia a 60-65 mm SL nei maschi e a 70-75 mm SL nelle femmine, e la deposizione delle uova avviene due volte l'anno durante aprile-maggio e luglio-settembre, o aprile-maggio e luglio-agosto (Hoda & Akhtar, 1985;
Hoda, 1986;
Hoda, 1987). In Kuwait, Clayton & Vaughan
(1988) riportano la stagione riproduttiva da marzo-aprile ad agosto, e la prima apparizione delle larve a metà luglio.
|
|
Note ecologiche:
Questa specie può raggiungere elevate densità (> 50 individui per 100 m²) sulle piane tidali fangose e sulle rive fangose di fiumi influenzati dalle maree, e si trova solitamente a livelli intertidali intermedi (Polgar et al., 2009;
Clayton & Vaughan, 1988;
Clayton, 1985). B. dussumieri
costruisce sistemi peculiari di cunicoli sotterranei e di territori poligonali circondati da mura di fango (vedi foto C-E) [Clayton & Vaughan, 1982; 1986 (B. boddaerti = B. dussumieri,
incorrettamente identificato); Clayton & Wright, 1989], che potrebbero avere la funzione di ridurre le interazioni aggressive intraspecifiche, o di facilitare la crescita delle diatomee bentiche (una sorta di "acquacoltura"), o entrambe [Clayton, 1987; quest'ultima ipotesi è stata supportata da Chen et al. (2009) per simili strutture realizzate da
B. pectinirostris in Cina]. L'etogramma è stato descritto da Clayton
& Vaughan (1988), che ha anche descritto una partizione spaziale fra individui territoriali di più grandi dimensioni e individui più piccoli non territoriali, o "erranti", che mostrano comportamenti gregari. Il comportamento termico e stagionale è stato descritto da Tytler & Vaughan (1983). Questi pesci sono predati da numerosi predatori, dagli uccelli ai serpenti ai coccodrilli (Clayton, 1988;
1993)
al centro: Qeshm Island, Hormozgan, Iran: una piana di fango densamente popolata (foto: E. Kamrani, 2007, con il permesso dell'autore)
|
|
|
|
Distribuzione:
Golfo Persico, Oman, Pakistan ed India occidentale; località tipo di B. dentatus: Bombay (= Mumbai), Maharashtra, west India; località tipo di B. chamiri: Stretto di Hormoz, Hormozgan, Iran (Murdy, 1989)
|
Note:
Valenciénnes (in Cuvier & Valenciènnes, 1837)
considerò B. dussumieri e B. dentatus come specie differenti.
Day (1876) discriminò B. dussumieri da B. dentatus (da popolazioni simpatriche in Bombay = Mumbai, India; e provincia del Sind e Karachi, Pakistan) dalla presenza di pinne pelviche più corte, una D1 più bassa, e dalla presenza di uno spazio interdorsale.
Secondo Berg (1949), le varie specie nominali furono basate su differenze ontogenetiche o sessuali: B. chamiri fu basato sui giovani, che hanno un numero inferiore di denti;
B. dussumieri fu basato sulle femmine (che hanno pinne dorsali più piccole); e B. dentatus sui maschi (citato da Brian Coad; visitato il 10/2009).
Hoda (1987) considerò B. dussumieri e B. dentatus specie separate, e addirittura descrisse diverse relazioni lunghezza peso e la crescita relativa di alcune parti del corpo; B. dentatus fu studiato anche da Mansuri et al. (1982) nel Gujarat, e da Soni e George
(1986). In seguito, Murdy
(1989) sinomizzò tutte queste specie.
Diversi autori hanno apparentemente confuso B. dussumieri dal Golfo Persico con B. boddarti,
che qui non è presente (e.g. Al-Kadhomiy & Hughes,
1988; Clayton & Vaughan, 1988); tuttavia, B. boddarti è presente lungo le coste occidentali dell'India
(e.g. Mumbai), dove queste due specie potrebbero ancora essere simpatriche (Murdy, 1989).
Mirza et al. (1996) hanno nuovamente discriminato B. dentatus da B. dussumieri
sulla base dell'altezza di D1, della profondità del corpo, del numero dei denti e sulla presenza/assenza di una connessione fra le pinne dorsali. Più recentemente, Abdoli et al. (2009) hanno trovato diversi esemplari di dimensioni inusualmente grandi in Iran meridionale (vedi foto A, B), che somigliano molto al B. dentatus di Valenciénnes (vedi tavola C); ho esaminato esemplari di dimensioni e forme simili nella collezione del Natural History Museum di Londra.
Polgar & Crosa (2009) hanno proposto che la specie malese descritta nello Stretto di Malacca da Takita et al. (1999) e Polgar & Khaironizam (2008) come B. dussumieri, sia in realtà B. pectinirostris.
|
Fotografie di Boleophthalmus dussumieri:
|
A: un esemplare di grandi dimensioni raccolto in Iran, Hormozgan, ad ovest di Bandar Abbas (la barra è lunga 10 mm; foto: G. Polgar, Università di Hormozgan, laboratorio di Biologia Marina, 2007); B: dettaglio della testa dell'esemplare in A: si notino le grandi papille dermiche, simili ad una peluria (foto: G. Polgar, 2007); C:
territori poligonali su di una piana tidale; alcuni ricercatori stanno campionando saltafango colpendoli con una canna, una tecnica tradizionale (foto: G. Polgar, Bandar Khamir, Hormozgan, Iran, 2007); D: un'altra foto dei territori poligonali di B. dussumieri (foto: E. Kamrani, Qeshm Island, Hormozgan, Iran, 2007)*; E: un'altra foto di territori poligonali (foto: D.A. Clayton, Sulaibikhat Bay, Kuwait, 1987); F-K: alcuni scatti di una sequenza aggressiva: i due pesci sono in formazione parallela o anti-parallela, o a T, con entrambe le pinne dorsali e la caudale erette (Clayton & Vaughan, 1988); G: una "promeade" in formazione parallela, con le bocche aperte (un P. waltoni è anche visibile sullo sfondo); H: un violento morso; I-K: un "colpo di coda", un comportamento non riportato dagli autori citati; L: a close-up in
aquarium (foto: G. Polgar, Università di Hormozgan, laboratorio di Biologia Marina, 2007) - * con il permesso dell'autore
|
Disegni di Boleophthalmus dussumieri:
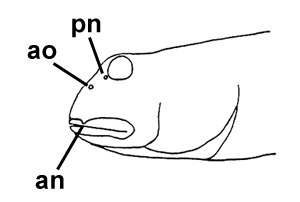
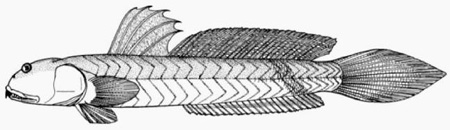
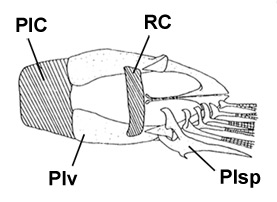
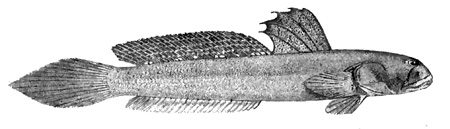
A: pori cefalici sensoriali e nasali di Boleophthalmus spp.: an = narice anteriore; ao = poro del canale oculoscapulare anteriore; pn = narice posteriore (modificato da Murdy, 1989); B: disegno di B. dussumieri di Susan Laurie-Bourque (Brian W. Coad's Personal Website)*; C: Boleophthalmus dussumieri Valenciénnes (sopra) e B. dentatus Valenciénnes (sotto) (Cuvier e Valenciénnes, 1837); D: vista ventrale del cinto pelvico di B. boddarti (gli elementi della pinna pelvica sono stati rimossi dal lato sinistro: PIC = cartilagine pelvica intercleitrale; Plv = pelvi; RC = cartilagine rettangolare; Plsp = spina della pinna pelvica (modificato da Murdy, 1989)*; E: Boleophthalmus dentatus Valenciénnes (Day, 1889) - * con il permesso dell'autore
|