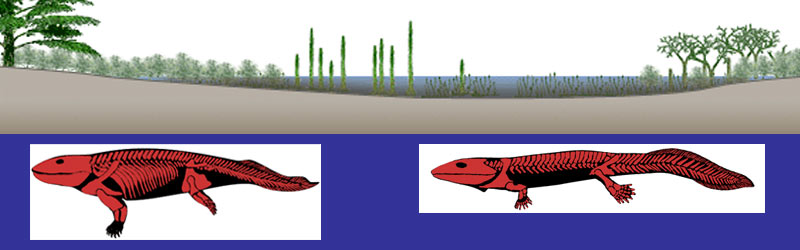
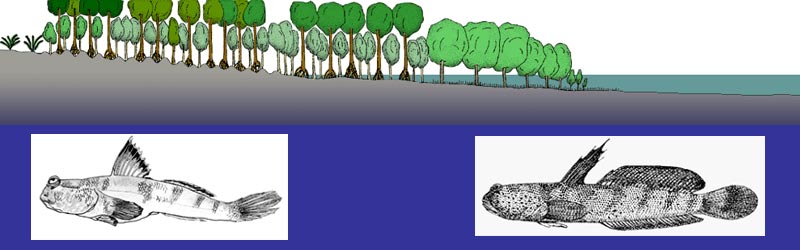
Is it possible to find any evolutionary convergent feature in these two independent pathways
to terrestriality?
Are at least some of the selective pressures operating on mudskippers similar to those ones that acted on aquatic tetrapodomorph sarcopterygians and on semi-aquatic early tetrapods 380-350 million years ago?
Could mudskippers teach us something about some of the evolutionary opportunities and challenges that our tetrapod ancestors faced
right "at the water's edge" in the Late Devonian?
The selective pressures that led vertebrates to the water-air transition, as well as those environmental conditions in which
this gradual evolutionary process took place, are both highly debated and subject to much speculation.
Palaeontological data are still quite fragmentary (Clack, 2002; see also The Devonian
opportunity).
Nonetheless, some studies suggest that this process took place in periodically flooded alluvial plains and/or in estuarine intertidal
wetlands, environments very similar to those experienced by mudskippers (Murphy, 2005; Schultze, 1999; Clack, 2002).
Furthermore, mudskippers may have remained in such environmental conditions throughout their evolutionary history
(see also Systematics & Biogeography ).
On the other hand, other scientists (Graham & Lee, 2004) agree for the most part with Romer's most widely accepted theory,
that the origin of tetrapods took place in tropical lowlands, seasonally dried up by chronical water shortages (Romer &
Parsons, 1986).
According to this theory, the first prototetrapods came out of water not to 'move onto land', but simply to avoid inhospitable aquatic
conditions, e.g. moving from a drying pool to a bigger one, not unlike several modern freshwater bimodal breathers.