|
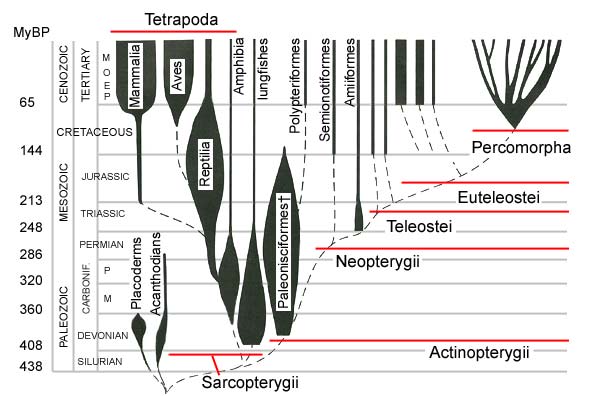
Mudskippers are currently included in the subfamily Oxudercinae (Teleostei, Acanthopterygii, Percomorpha, Gobiiformes, Gobioidei, Gobiidae; Nelson, 2006; Thacker et al., 2015), whose diagnostic characters were defined by Hoese (1984).
Murdy (1989) proposed a complete cladistic revision of this group. This revision was very welcome to mudskipper taxonomy, which has been intricate and chaotic ever since the 17th century.
In particular, Murdy described 13 morphospecies of Periophthalmus. Five new species of the genus Periophthalmus were subsequently
described (P. magnuspinnatus Lee et al., 1995, P. spilotus Murdy & Takita, 1999; P. walailakae Darumas & Tantichodok, 2002; P. darwini Larson & Takita, 2004; P. takita Jafaar & Larson 2008), while P. novaeguineaensis Eggert was redescribed (Jafaar & Larson 2008), and P. variabilis Eggert restored (Jafaar et al. 2009). One species of Boleophthalmus was also described after Murdy (1989): Boleophthalmus poti Polgar et al., 2013. Therefore, presently oxudercines include 41 morphospecies and 10 genera:
Apocryptes Valenciennes, 1837* (1)
Apocryptodon Bleeker, 1874 (2)
Boleophthalmus Valenciennes, 1837* (6)
Oxuderces Eydoux & Souleyet, 1850 (2)
Parapocryptes (Valenciennes, 1837)* (2)
Periophthalmus Bloch & Schneider, 1801 (18)
Periophthalmodon Bleeker, 1874 (3)
Pseudapocryptes Bleeker, 1874 (2)
Scartelaos Swainson, 1839 (4)
Zappa Murdy, 1989 (1)
*In: Cuvier & Valenciennes, 1837
The monophyly of Oxudercinae has been recently questioned by several molecular
studies (Agorreta & Rüber, 2012;
Agorreta et al., 2013;
Thacker, 2003;
Tornabene et al., 2013). In particular, oxudercine gobies appear
paraphyletic relative to amblyopines (subfamily Amblyopinae, or worm-eel gobies, which appear as polyphyletic).
Periophthalmus also appears as the sister group of all other oxudercine and amblyopine genera so far examined.
Consistently, a molecular systematic and phylogeographic study (Polgar et al., 2014) showed that this genus has a long evolutionary history (>30 million years) in coastal wetland ecosystems.
The vernacular term 'mudskippers' implies the presence of an amphibious lifestyle. For this reason, 'mudskippers' should be used to indicate those oxudercine species that "are fully terrestrial for some portion of the daily cycle"
(Murdy, 1989). Such species are included in the genera Boleophthalmus, Periophthalmodon,
Periophthalmus, and Scartelaos. Based on the observations by Polgar et al.
(2010) on Zappa confluentus, this latter species should be included in this definition.
|
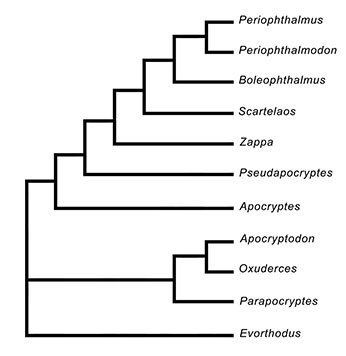
Cladogram of the genera of Oxudercinae (Maximum Parsimony tree), based on morphological, osteological, and eco-ethological characters; the gobionelline goby Evorthodus lyricus was used as outgroup (Murdy, 1989). The original dataset was analysed in PAUP v.4 (Swofford, 2002). With permission from the author
|