|
Nomi comuni:
Australia |
inglese |
silver-lined mudskipper |
China |
cinese mandarino |
银线弹涂鱼, 銀線彈塗魚 (yin xian tan tu yu - saltafango striato d'argento) |
Danimarca |
danese |
båndet dyndspringer |
Guam |
inglese |
barred mudskipper |
Indonesia |
javanese |
blodok |
Italia |
italiano |
perioftalmo argentato* |
Giappone |
giapponese |
minami tobihaze |
Gran Bretagna |
inglese |
barred mudskipper, silver-lined mudskipper |
Micronesia |
inglese |
barred mudskipper |
Nuova Caledonia |
kumak |
bwaxit |
Papua Nuova Guinea |
inglese |
barred mudskipper |
Filippine |
tagalog |
bia |
Isole Solomone |
gela |
iga kukulu mana |
Isole Solomone |
inglese |
silver-lined mudskipper |
Sud Africa |
inglese |
bigfin mudhopper |
Sud Africa |
afrikaans |
Grootvin-modderspringer |
* nome proposto
|



Periophthalmus argentilineatus. Morib, Malaysia peninsulare;
vista laterale (sopra); vista dorsale (al centro); vista ventrale (sotto);
la barra è lunga 10 mm - esemplare appena morto (foto: G. Polgar, 2005)



Periophthalmus argentilineatus.
RMNH 4593, coll. Bleeker 1879
possibile sintipo (secondo Murdy, 1989) di Periophthalmus dipus Bleeker, 1854
Sumatra, Java, Flores (Eggert, 1935)
vista laterale (sopra); vista dorsale (al centro); vista ventrale (sotto);
la barra è lunga 10 mm - esemplare fissato e conservato in etanolo, maschio
(foto: G. Polgar, 2007)
|
|
Sinonimi:
|
Periophthalmus argentilineatus |
Valenciennes, 1837* |
(sin. senior, combinazione originale) |
|
Periophthalmus dipus |
Bleeker, 1854 |
(sinonimo junior) |
Periophtalmus koelreuteri argentilineata |
Bianconi, 1857 |
(invalido, errore sintattico) |
|
Euchoristopus kalolo regius |
Whitley, 1931 |
(sinonimo junior) |
|
Periophthalmus argentilineatus argentilineatus |
Valenciennes, 1837* |
(sinonimo senior)** |
|
Periophthalmus argentilineatus striopunctatus |
Eggert, 1935 |
(sinonimo junior) |
|
Periophthalmus dipus angustiformis |
Eggert, 1935 |
(sinonimo junior) |
|
Periophthalmus dipus dipus |
Eggert, 1935 |
(sinonimo junior)** |
|
Periophthalmus dipus parvus |
Eggert, 1935 |
(sinonimo junior) |
|
Periophthalmus sobrinus |
Eggert, 1935 |
(sinonimo junior) |
|
Periophthalmus vulgaris ceylonensis |
Eggert, 1935 |
(sinonimo junior) |
|
Periophthalmus vulgaris notatus |
Eggert, 1935 |
(sinonimo junior) |
|
Periophthalmus vulgaris regius |
Whitley, 1931 |
(sinonimo junior)** |
|
Periophthalmus vulgaris vulgaris |
Eggert, 1935 |
(sinonimo junior) |
*In: Cuvier & Valenciennes, 1837
**sensu Eggert (1935)
Catalogo del materiale tipo (ultimo aggiornamento: 15/08/2007)
Etimologia:
'Periophthalmus' è un nome composto dal greco 'peri' (attorno), e 'ophthalmôn' (occhio), riferendosi all'ampio campo visivo di queste specie
'argentilineatus' significa 'striato d'argento' in latino, riferendosi alle striature verticali argentate sui fianchi (Murdy, 1989)
|
Lunghezza massima registrata:
93 mm SL (Murdy, 1989)
Colorazione in vivo (Murdy, 1989; Takita et al., 2011; oss. pers.:
Malaysia peninsulare, Nord Sulawesi, Australia NT):
colorazione di fondo da marrone chiaro a grigio scura su dorso e fianchi, ventralmente biancastra; testa ventralmente bianca; molti piccoli punti bianchi su guance e opercoli; cospicue striature verticali argentate sui fianchi, più evidenti
nella zona ventrale; 3-8 barre dorsali a sella marrone scuro irregolari possono essere visibili; D1 con sfondo da rossastro a marrone, una larga striscia inframarginale da nera a marrone scuro
(che può essere più scura nella porzione anteriore, per es. foto H, L), e molti piccoli punti bianchi al
di sotto; margine da bianco a trasparente. D2 con una striscia nera in posizione mediana
bordata da due strisce bianche in alcune varianti; chiazze irregolari marroni in posizione basale e prossimale su
elementi e membrana (che in alcune varianti possono fondersi fino a formare uno sfondo marrone con pochi o nessun punto
bianco), e margine da giallo a rosso; pinna caudale grigiastra, con serie di punti scuri da grigi a marroni a rossastri lungo i raggi; pinna anale
ialina, in alcuni esemplari distalmente da grigiastra a arancione; pinne pettorali grigiastre; pinne pelviche
più scure in posizione prossimale, ventralmente biancastre, e da biancastre a giallastre in posizione dorsale
Colorazione in liquido conservante
(Murdy, 1989; oss. pers.):
colorazione di fondo dorsalmente e lateralmente da grigia a marrone scuro, da biancastra a grigia sul ventre; alcuni punti biancastri possono rimanere visibili in alcuni esemplari; le strisce verticali argentee sui fianchi sono più visibili in posizione ventrale; il pattern dorsale delle bande a sella scure ed irregolari è frequentemente visibile; D1 con sfondo da marroncino a grigio ardesia, con molti punti bianchi sparsi in posizione prossimale ed una cospicua banda inframarginale nerastra; margine dal chiaro a trasparente;
D2 marroncina con una striscia da nera a marrone scuro in posizione mediana, e punti bianchi sparsi sulla membrana in posizione prossimale; pinna caudale grigiastra con serie di punti scuri sui raggi; pinne pettorali grigiastre; pinne pelviche
ventralmente bianche, grigiastre in posizione dorsale; pinna anale bianca
Diagnosi (Murdy,
1989):
D1 XI-XVI; elementi totali di D2 10-13; elementi totali della pinna anale 9-12; TRDB 18-26; conta longitudinale delle scaglie 64-100; larghezza della testa 14.3-22.6%SL; lunghezza delle pinne pelviche 11.3-15.2%SL; lunghezza della base della pinna anale 14.0-19.4%SL; lunghezza della base di D2 17.6-23.7%SL; pinne pelviche da quasi completamente a completamente separate, frenulo pelvico assente; margine di D1 concavo o diritto; pinne dorsali non connesse da membrana; D1 con una striscia inframarginale scura e numerosi punti bianchi almeno nella porzione prossimale; spine non allungate.
Almeno in alcune popolazioni dell'Africa orientale, il margine rossastro di D2 è assente nelle femmine adulte riproduttive (Magnus, 1981).
Vedi le note su P. kalolo sulla discriminazione fra queste due specie.
Il genere non è tuttora definito da sinapomorfie
Dieta:
carnivoro opportunista, apparentemente solo limitato dalle dimensioni della bocca (insetti, crustacei, uova di pesci, policheti, ecc.: Milward, 1974); tuttavia, differenti regimi alimentari ed una differente selettività sono state riscontrate nei due sessi, suggerendo la presenza di diverse strategie riproduttive legate al ciclo tidale sinodico; questi studi hanno anche messo in evidenza che almeno le popolazioni africane di questa specie si nutrono sia di notte che di giorno (Colombini et al., 1996). Stebbins & Kalk, 1961 e Macnae & Kalk, 1962 hanno studiato il contenuto stomacale di P. sobrinus (= P. argentilineatus) in Madagascar, trovando crostacei, insetti e policheti; tuttavia è probabile che non abbiano discriminato fra questa specie e P. kalolo, che in questa regione sono simpatriche (Clayton, 1993)
|
|
Riproduzione:
i maschi scavano tane riproduttive al di sotto del livello di MHWN (Mean High Water Neap - livello medio dell'acqua alta durante
le maree morte, davanti alle prime file degli alberi del mangrovieto), in aree non coperte da vegetazione, saltando con le pinne
spiegate per attirare le femmine e quindi indurle a deporre all'interno della tana; durante il breve periodo di coabitazione
hanno luogo diversi eventi di deposizione; in seguito solo il maschio rimane a fare la guardia al nido,
rimanendo nelle immediate vicinanze alla bassa marea e mantenendo una fase aerea nella camera delle uova; durante l'alta marea,
quando il nido viene sommerso, il maschio si rifugia al suo interno (Brillet, 1969a;
1970; 1975; 1976; 1980;
Magnus, 1981)
a sinistra: un maschio durante una fase della sequenza di corteggiamento (sventolamento della coda)
a destra: un esemplare ingoia una bolla d'aria prima di portarla all'interno della tana
(foto: a sinistra: Y. Ikebe, Giappone meridionale (© Mudskipper World, 2008); a
destra: fotogramma tratto da Magnus, 1981)
|
|
|
|
|
Note ecologiche (oss. pers.: Malaysia peninsulare, Nord Sulawesi, Australia NT):
localmente comune sulla piana tidale superiore alla bassa marea, di solito a distanze inferiori ai 2 m dall'acqua, mai distante dalla vegetazione a mangrovie, tipicamente lungo le rive delle insenature tidali e nei pressi delle pozze tidali in foreste a mangrovie pioniere.
Con la marea calante
P. argentilineatus segue il bordo dell'acqua per andare a nutrirsi sulla piana fangosa superiore o sulle rive fangose delle vie d'acqua tidali (Brillet, 1975; Magnus, 1981; Takita et al., 2011; oss. pers.); durante la fase di flusso i pesci seguono nuovamente la marea, rimanendo fuori dall'acqua: possono entrare in tane sotterranee, arrampicarsi su radici e tronchi (come P. chrysospilos), o attendere lungo il bordo dell'acqua, con le teste dirette verso l'acqua (Magnus, 1981). I dettagli potrebbero comunque essere più complessi, essendo in relazione sia con il ciclo tidale sinodico che con il ciclo giorno-notte (Colombini et al., 1995)
a sinistra: riva di una barra tidale con vegetazione a mangrovie ripariale (Avicennia sp.), un tipico habitat dove si può trovare P. argentilineatus alla bassa marea (Likupang, North Sulawesi, Indonesia)
a destra: un P. argentilineatus arrampicato su un tronco di Avicennia alba, in attesa della bassa marea (Tg. Piai, Malaysia peninsulare)
(foto di G. Polgar; a sinistra: 2005; a destra: 2006)
|
|
|
|
|
Distribuzione (Murdy, 1989):
dal Mar Rosso e costa orientale dell'Africa, verso est fino al Giappone meridionale, Australasia ed Oceania, fino alle isole Samoa;
località tipo: Irian Jaya e Molucche, Indonesia.
Si tratta della distribuzione più ampia fra tutti i gobidi oxudercini
|
|
|
Note:
Vedi Periophthalmus kalolo.
Analisi filogenetiche molecolari suggeriscono che questa specie includa almeno tre specie criptiche (Polgar et al., 2014), una morfologicamente compatibile con P. sobrinus Eggert (oss. pers.; distribuita nel Mar Rosso e in Africa orientale), una compatibile con P. vulgaris Eggert (oss. pers.; distribuita dallo Sri Lanka verso est fino alla Sumatra occidentale, Arcipelago della Sunda, Sulawesi, Filippine, Molucche, Papua Occidentale e Australia settentrionale); e una compatibile con P. argentilineatus Valenciennes (oss. pers.;
distribuita dalla Sumatra occidentale, verso est nell'Arcipelago della Sunda, Borneo sudorientale e Molucche)
|
Fotografie di Periophthalmus argentilineatus:
|
|
A: diversi individui cercano rifugio su una roccia emersa durante l'alta marea: in queste condizioni l'aggressività intraspecifica viene inibita (foto: Y. Yoshida, Baia di Isahaya, Giappone © isahayawan); B: P. argentilineatus mentre salta per fuggire da un pericolo: si notino le pinne chiuse ed aderenti al corpo;
C: un individuo con un granchio in bocca minaccia un conspecifico erigendo la prima pinna dorsale e scurendo il muso; D: diversi esemplari si arrampicano sulla mano del fotografo (foto B-D: Akinori Kamiya "Yamaneko", torrente tidale Ohashi, Giappone, 2001 © umisuzume, 2007)*; E: esemplare fijiano morto da poco (foto: J. Randall, Isole Fiji, 1973; fishbase, 2007); F: esemplare indonesiano morto da poco (foto: J. Randall, Baya di Ambon, 1975; fishbase, 2007); G: un altro esemplare indonesiano morto da poco (foto: G. Polgar, Nord Sulawesi, 2005); H: esemplare australiano morto da poco (foto G. Polgar, Vestey's Creek, Darwin, Northern Territory, 2007); I: un individuo in acquario minaccia la mano dell'allevatore; J: vista ventrale delle pinne pelviche (foto I, J: M. Tanabe, © Marli's mudskipperland, 2007);
K: un esemplare australiano alla bassa marea (foto: G. Polgar, Doctor's Gully, Darwin, Northern Territory); L: due maschi in confronto aggressivo (foto L. Mráz, South Africa, St. Lucia estuary, cape Vidal, Sugar Loaf, 2005 © naturephoto-CZ)* - * con il permesso dell'autore
|
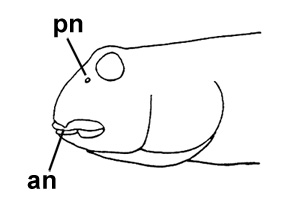
Disegni di Periophthalmus argentilineatus:
A: pori cefalici sensoriali e nasali di
Periophthalmus spp.: an = narice anteriore; pn =
narice posteriore (modificato
da Murdy, 1989)*; B: Periophthalmus vulgaris regius Eggert, 1935
(Eggert, 1935); C:
Euchoristopus dipus (Bleeker) (Bleeker, 1983)*;
D: Euchoristopus argentilineatus (Valenciénnes) (Bleeker, 1983)*
- * con il permesso dell'autore
|