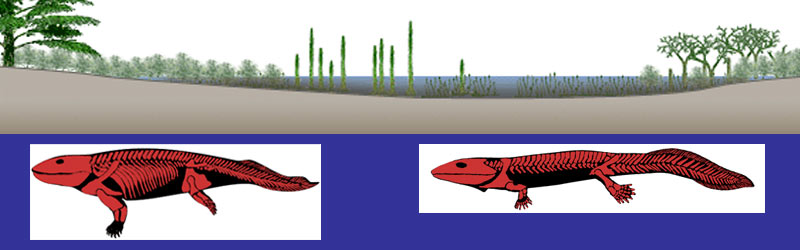
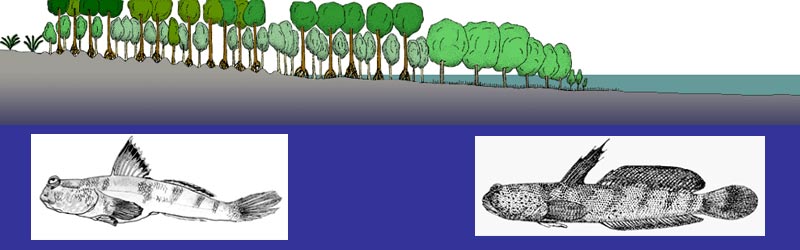
E’ possibile trovare tratti convergenti in questi due indipendenti percorsi evolutivi verso la terrestrialità?
E’ possibile comparare almeno alcune delle pressioni selettive che stanno operando sui saltafango oggi con quelle che agirono sui tetrapodomorfi acquatici e sui primi tetrapodi semi-acquatici 380-350 milioni di anni fa?
E’ possibile imparare dai saltafango quali furono le opportunità evolutive e le sfide che i nostri antenati tetrapodi affrontarono 'lungo il bordo dell’acqua' nel devoniano superiore?
Le pressioni selettive che condussero i vertebrati alla transizione acqua-terra, così come quelle condizioni ambientali in cui questo graduale processo evolutivo ebbe luogo, sono a tutt’oggi argomenti perlopiù speculativi, oggetto di un intenso dibattito scientifico.
I dati paleontologici infatti sono ancora frammentari (Clack, 2002; vedi anche L’opportunità del devoniano).
Alcuni studi tuttavia suggeriscono che questo processo abbia avuto luogo in piane alluvionali inondate periodicamente e/o in aree intertidali ad estuario: ambienti molto simili a quelli in cui vivono oggi i saltafango (Murphy, 2005; Schultze, 1999; Clack, 2002).
I saltafango inoltre, potrebbero essere sempre rimasti in tali condizioni ambientali nel corso della loro breve storia evolutiva (vedi anche Sistematica e biogeografia).
D’altro canto, la teoria più largamente condivisa (Graham & Lee, 2004) è quella di Romer, secondo cui i tetrapodi ebbero origine in aree alluvionali tropicali a siccità stagionale (Romer & Parsons, 1986).
Secondo questa teoria, i primi prototetrapodi uscirono fuori dall’acqua non per muoversi 'verso condizioni più terrestri', ma per evitare condizioni acquatiche inospitali, come ad esempio per spostarsi da una pozza quasi prosciugata o affollata, verso una pozza più grande; comportamenti simili si possono osservare in diverse specie di pesci dulcacquicoli tropicali capaci di respirazione bimodale.