Ricerca personalizzata

Ultimo aggiornamento: 14/07/2007
|
Ricerca personalizzata
|
Ultimo aggiornamento: 14/07/2007 |
|
Diversi cambiamenti ecologici hanno giocato un ruolo determinante nella transizione evolutiva acqua-terra dei tetrapodi. |
 | |
|
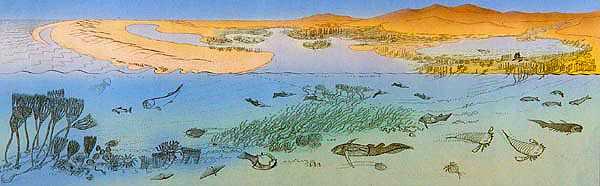
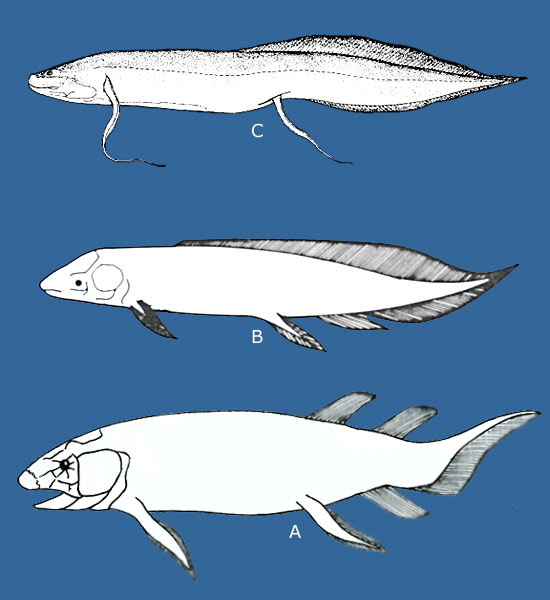
A sinistra: ricostruzione di un tipico paesaggio del devoniano inferiore; a destra: due tetrapodi primitivi lungo le sponde di un corso d’acqua del devoniano superiore: Acanthostega gunneri (in basso) ed Ichthyostega stensioei (in alto). | ||
|
|
|
|
|
| |
|
|
|
|
|
|
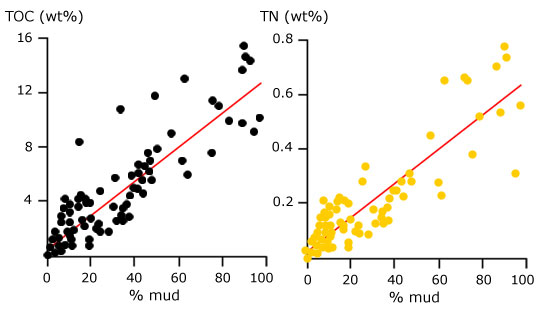
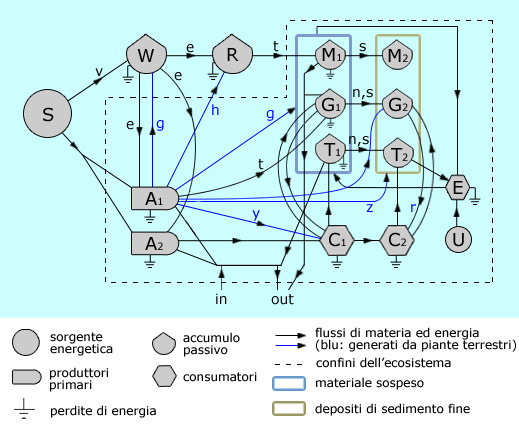
Inoltre, la vegetazione in rapida espansione doveva esercitare nelle acque basse una forza di attrito che diminuiva fortemente l’energia deposizionale (come oggi avviene nelle foreste a mangrovie: Mazda et al., 1997a,b), aumentando così il tasso di sedimentazione del sedimento fine. |
  | |
|
|
 |
| |
|
|
|
|
|
|
|
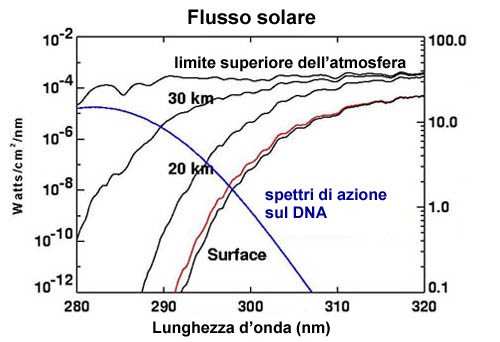 |
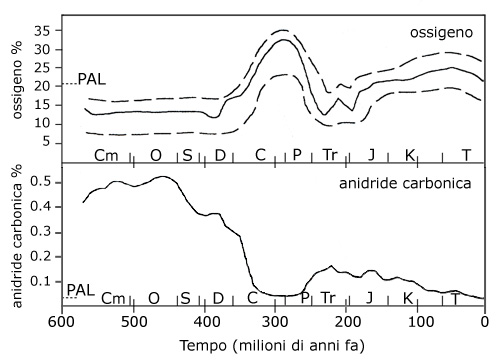
L’aumento dei livelli di ossigeno nell’atmosfera rese anche possibile l’ispessimento dello strato di ozono devoniano, già formatosi in seguito all’azione fotosintetica degli autotrofi unicellulari durante il proterozoico (2500-540 milioni di anni fa: Bullini et al., 1998). | |
|
|
|
Quali furono gli effetti di questi drammatici cambiamenti sulle condizioni di questi ambienti di acque basse? Abbondanti risorse trofiche, ambientali e metaboliche, così come habitat liberi da competitori vertebrati, erano a disposizione di qualsiasi linea evolutiva di pesci che fosse riuscita ad aumentare gradualmente il proprio grado di terrestrialità (Clack, 2002). |
| |
|
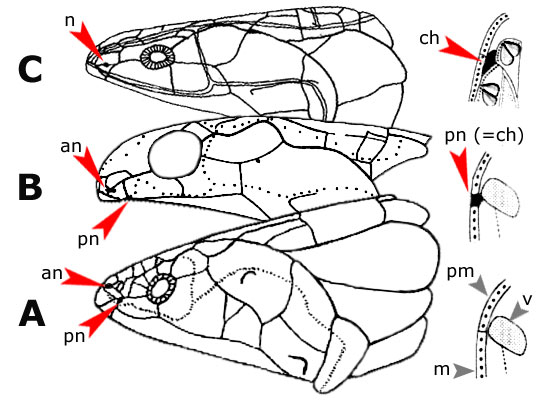
Evoluzione delle coane nei tetrapodomorfi. |

Tutto il contenuto su questo sito web (inclusi testi, fotografie e qualsiasi altro lavoro originale), se non altrimenti specificato, è tutelato da una licenza del tipo
Creative Commons License