
Ricerca personalizzata

Ultimo aggiornamento: 16/04/2010
|
Ricerca personalizzata
|
Ultimo aggiornamento: 16/04/2010 |
|
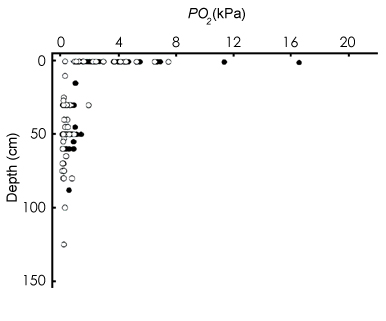
In molti pesci che respirano ossigeno atmosferico, come nei gobidi oxudercini, la capacità di respirare sia in acqua che in aria (respirazione bimodale) sembra essere un adattamento a basse concentrazioni ambientali di ossigeno (ipossia) ed alte concentrazioni di anidride carbonica (ipercarbia o ipercapnia) (Graham, 1997; Horn et al., 1999; Ultsch, 1996). Tali adattamenti consentono a questi pesci di vivere in acque a forte inquinamento organico. |
|
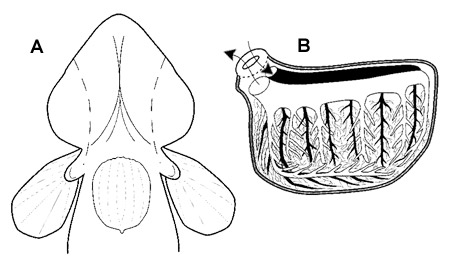 Organi per la respirazione aerea ('Air Breathing Organs': ABOs) di Pseudapocryptes lanceolatus. A: vista ventrale della testa, pinne pettorali e pelviche; le linee tratteggiate indicano l’estensione delle tasche opercolari espanse. B: vista sagittale del sistema circolatorio opercolare (le frecce indicano la direzione del flusso sanguigno). Modificato da Graham, 1997, con il permesso della Elsevier. |
|
 |
|
|
|
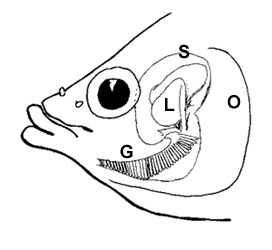
Diverse specie di gobidi possono tenere bolle d’aria nel cavo orale mentre sono in acqua (AG= 'air gulping': inghiottire aria; Gee & Gee, 1995). Le specie di saltafango più terrestri (Periophthalmus e Periophthalmodon spp.) lo fanno anche quando emergono, sigillando le camere opercolari per mezzo di una specifica valvola ventro-mediale (Sponder & Lauder, 1981; Clayton, 1993; Martin & Bridges, 1999; Graham, 1997). Fuori dall’acqua queste specie respirano attraverso la cute e le mucose del cavo oro-bucco-faringeo; in acqua, dove l’assorbimento dell’ossigeno è meno efficente, fanno uso sia della respirazione cutanea che di quella branchiale (Clayton, 1993; Ishimatsu et al., 1999; Takeda et al., 1999). |
|
|

Tutto il contenuto su questo sito web (inclusi testi, fotografie e qualsiasi altro lavoro originale), se non altrimenti specificato, è tutelato da una licenza del tipo
Creative Commons License

