Common names:
China |
Chinese Mandarin |
许氏齿弹涂鱼, 許氏齒彈塗魚 (Xu shi chi tan tu yu - Xu’s teethed mudskipper) |
Denmark |
Danish |
stor dyndspringer |
Philippines |
Tagalog |
bia |
Finland |
Finnish |
helmiliejuryömijä |
Germany |
German |
Schlammspringer |
Indonesia |
Javanese |
blodok, gabus lawut |
Italy |
Italian
|
saltafango di Schlosser* |
Malaysia |
Bahasa Malaysia |
tembakul, ikan belacak gergasi |
Viet Nam |
Vietnamese |
cá thòi lòi sơ - lốt - sơ |
United Kingdom |
English |
giant mudskipper |
* proposed name
|
|
|
Synonyms:
Etymology:
'Periophthalmodon' is a compound form from 'Periophthalmus' and the Greek 'odous' (tooth), which refers to the prominent teeth and strong similarity to the genus Periophthalmus
the species is named after J.A. Schlosser, who collected the material for the original
description (Murdy, 1989)
|
Maximum recorded length:
240 mm SL (Swennen et al., 1995)
Live colouration (Murdy, 1989; pers. obs.: Peninsular Malaysia):
ground colour dorsally and laterally dark to pale brown,
ventrally whitish to grey; frequently visible a typical black stripe
coursing from eye posteriorly across dorsal opercular edge,
continuing dorsal to pectoral fin, and up to caudal peduncle;
6-8 dark brown saddle-like diagonal bars visible in dark light,
especially at night; snout, cheeks
and flanks with whitish to pale blue, iridescent speckles, as
big as the underlying scale; D1 reddish brown to dark brown with
white margin; D2 brown with white margin, with a darker stripe
inframarginally; caudal fin dark brown; anal fin hyaline; pectoral
fins brownish; pelvic fins hyaline
Colouration on preservation
(Murdy, 1989; pers. obs.: Peninsular Malaysia):
ground colour brown to slate grey dorsally and laterally, whitish to
yellowish ventrally; frequently visible the dorsal dark stripe from eye
to caudal region; saddle bars rarely retained; white speckles on snout,
cheecks and flanks may be retained; D1 and D2 dark brown with pale to
translucent margins; inframarginal stripe in D2 may be retained; caudal fin
dark brown to greyish; anal fin whitish to yellowish; pectoral fin brown to
greyish; pelvic fin whitish to yellowish
Diagnosis (Murdy, 1989):
D1 VI-IX; pectoral fin rays 16-19;
length of D1 base 10.3-16.7%SL; when visible, a narrow
dark stripe courses from eye up to caudal peduncle; isthmus lacking scales; snout scaled; dorsal fins never contiguous;
pelvic fins completely fused and with prominent pelvic frenum.
The genus is characterised by the presence of two rows of teeth on the upper jaw;
in this species the second row of teeth in the upper jaw
contains few and smaller teeth, often half buried in the mucosa (pers. obs.)
Diet:
carnivorous; it mainly feeds on crabs (Khoo, 1966,
cited in Clayton, 1993;
Macintosh, 1979;
Sasekumar et al., 1984), but can prey also on
other locally abundant animals (Lim, 1971,
cited in Clayton, 1993), included
syntopic species of mudskippers (e.g.
Boleophthalmus boddarti: pers. obs.: Peninsular Malaysia)
Reproduction:
the male and female cohabit inside the burrow for a period, maintaining the
burrow and leaving it at ebb and flood tide to feed or take shelter at the water's
edge, respectively (pers. obs.). Mazlan & Rohaya (2008) measured gonadosomatic and hepatosomatic indexes of a Malayan population; the inferred breeding season was from June to October, during the southwest monsoon; these estimates indicate a size at first maturity of 12-14 cm TL (females); and a maximum length of 29 cm TL. The first developmental stages and larval morphology were partly described by Tsuhako et al. (2003)
|
|
Ecological notes:
during low tide Pn. schlosseri occurs on open mudflats, along mangrove forests' marine fringes, along creeks' banks, and even inside mangals: never far from the water's
edge or from the large pools of their burrows; larger and reproductive individuals dig deep tunnels in the soft mud: inside mangals burrows are only present on the beds of
ephemeral inlets (Takita et al., 1999; Polgar & Crosa, 2009); smaller individuals are frequently found along the banks of ephemeral inlets inside mangrove forests (pers. obs.: Peninsular Malaysia)
left: Tanjung Piai, Peninsular Malaysia: mud flat; this area is
regularly crossed by large moving individuals of Pn. schlosseri during ebb and flood tide and
burrows are abundant
right: Kukup Is., Peninsular Malaysia: bed of an ephemeral inlet in
the high mangrove forest at low tide; note the absence of tree plants and the large circular openings of Pn. schlosseri's burrows
both are typical examples of the habitat of this species (photo: G. Polgar, 2006)
|
|
 |
|
|
Distribution:
South East Asia; type locality: Ambon, Indonesia (Murdy, 1989)
|
Remarks:
Pallas (1770) described Gobius schlosseri from Ambon (Indonesia), which is out of the known distributional range of Periophthalmodon schlosseri (Pallas), and within the distributional range of Periophthalmodon freycineti (Quoy & Gaimard); no type material is presenlty available. Later on, G. schlosseri Pallas was included in the genus Periophthalmus, established by Bloch & Schneider (1801); it is worth noting that in this same work these latter authors described the species Periophthalmus ruber, that was later on synonymised with Periophthalmodon schlosseri (Pallas) by several authors (Eggert, 1935; Koumans, 1953; Murdy, 1989). The holotype of Periophthalmus ruber Bloch & Schneider (ZMB 2143) is available; although in poor conditions, the height of D2, the length of the interdorsal gap, and the size of D1 would suggest that this specimen is instead Periophthalmodon freycineti (Quoy & Gaimard) (see photo above); it is worth noting that Bloch & Schneider (1801) reported for P. ruber a D1 count of 12 elements, which is neither consistent with Pn. schlosseri, nor with Pn. freycineti. Also the type locality of P. ruber (Tranquebar, India) is inconsistent with the known distributional range of both species; nonetheless, several authors agree that the type localities of Bloch & Schneider (1801) are often unreliable (Cuvier & Valenciénnes, 1837; Murdy, 1989; see also P. barbarus). Periophthalmus schlosseri (Pallas) became the type species of the genus Periophthalmodon after Bleeker (1874), based on the double row of teeth on the upper jaw of these species.
Pn. schlosseri is traditionally consumed in Peninsular Malaysia by native people (Orang Laut, or "sea-gypsies"; pers. obs.)
|
Photographs of Periophthalmodon schlosseri:
|
A: Pn. schlosseri at night: note the banded pattern of the
colouration (photo: G. Polgar, Kukup jetty, Peninsular Malaysia, 2006); B: the typical trackway
of a Pn. schlosseri (photo: G. Polgar, Carey Is., Peninsular Malaysia, 2006); C:
an individual at flood tide, swimming at the water surface (photo: G. Polgar, Carey Is., Peninsular Malaysia,
2006); D: inside its burrow on the forest floor: note the wide main opening (photo: G. Polgar, Kukup Is.,
Peninsular Malaysia,
2006); E: a juvenile in a shallow pool nearby an inlet inside the forest, at low tide: the specimen is nearby some pencil-like
pneumatophores of Avicennia sp., with a diameter of about 1 cm (photo: G. Polgar, Kukup Is., Peninsular Malaysia,
2006); F: two large specimens fighting for an emerged palm frond during high tide (photo: Z. Pogonyi, Pulau Ubin, Singapore, 2006)*;
G: a juvenile in a plastic bag (photo: G. Polgar, Kukup Is., Peninsular Malaysia, 2006); H:
another large individual on the mudflat at low tide: note how the lateral stripes are not visible (photo: G. Polgar, Tg. Piai, Peninsular Malaysia, 2007); I:
an individual in aquarium on a dark background: note the banded pattern (photo: G. Polgar, Kuala Lumpur,
Peninsular Malaysia, 2006); J: the same specimen photographed in I on a white
background: the banded pattern is absent (photo: G. Polgar, Kuala Lumpur, Peninsular Malaysia, 2006); K: spitting mud out of the burrow during maintenance (photo: G. Polgar, Tg. Piai, Peninsular Malaysia, 2007); L: Pn. schlosseri cooked in different ways by the Orang Laut fishermen: steamed
with ginger; fried; and in a soup, as fish balls (photo: G. Polgar, Kukup town, Peninsular Malaysia, 2006) - * with permission
|
Drawings of Periophthalmodon schlosseri:
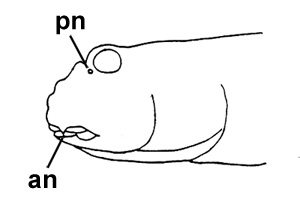
A: cephalic sensory and nasal pores of Periophthalmodon spp.: an = anterior
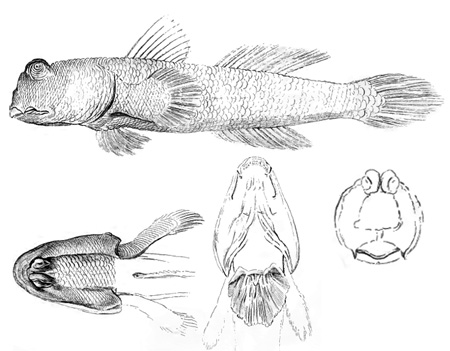
nostril; pn = posterior nostril (modified from Murdy, 1989)*; B: Periophthalmodon schlosseri (Pallas) (Rainboth, 1996); C: Gobius schlosseri Pallas (modified from Pallas, 1770); D: Periopthalmodon schlosseri (Pallas) (Bleeker, 1983)* - * with permission
|