Common names:
Bangladesh |
Bengali |
dahuk |
Bangladesh |
English |
goggle-eyed goby |
China |
Chinese Mandarin |
薄氏大弹涂鱼, 薄氏大彈塗魚 (Bo shi da tan tu yu - Bo’s great mudskipper) |
Philippines |
Tagalog |
bia |
Finland |
Finnish |
nuoliliejuryömijä |
Germany |
German |
Glotzauge |
Japan |
Japanese |
mutsugoro |
Indonesia |
Javanese |
blodok, lunjat |
Italy |
Italian |
boleoftalmo di Boddart* |
Malaysia |
Bahasa Malaysia |
ikan belacak binte biru |
Thailand |
Thai |
บู่ลาย, bulai |
Viet Nam |
Vietnamese |
cá bong sao, cá lác bo đa, cá thòi lòi |
* proposed name
|
|
|
Synonyms:
Etymology:
'Boleophthalmus' depicts the capability of these fishes to rapidly raise their eyes above the level of their orbits, as if they were ejected out of their orbits: from 'bolê' (ejected), and 'ophthalmôn' (eye) (Cuvier & Valenciénnes, 1837)
the specific name is after Pierre Boddaërt, who collected the material for the original description (Pallas, 1770); 'boddaerti' is an unjustified emendation (emendatio incorrecta) of the original spelling (Murdy, 1989; Catalog of Eschmeyer)
|
Maximum recorded length:
135 mm SL (Murdy, 1989)
220 mm TL (Kottelat et al., 1993)
Live colouration (Murdy, 1989; Takita et al., 1999; Polgar & Crosa, 2009):
ground colour dorsally and laterally brown to greenish, ventrally whitish to grey, darker behind anus and under the head; head and trunk
with bluish, iridescent small speckles; dark to black blotches on head and nape; a dark stripe may be present
from anterior nostril to dorsal tip of operculum; 7-8 diagonal saddle-like dark brown to black blotches on dorsum, posteriorly usually
extending below lateral midline; D1 greenish with numerous small pale blue speckles, distally with paler blotches (diagnostic) behind the first 2-3- spines; D1 yellow in young and juveniles;
D2 membrane greenish, with series of bluish spots between rays;
caudal fin grey to blue;
anal fin hyaline with an inframarginal dark stripe, to completely blackened; pectoral fins with orange to yellow membrane, dorsally with a
black to dark brown margin (diagnostic); muscular base of pectoral fins with numerous scattered whitish to bluish speckles; pelvic fins dorsally partially
pigmented, ventrally whitish and pigmented proximally
Colouration on preservation (Murdy, 1989;
pers. obs.: Bangladesh, Peninsular Malaysia, India, Indonesia):
ground colour dark brown to dark grey dorsally and laterally, whitish to greyish ventrally, with dark grey to bluish lips and chin; saddle
bars and dark blotches on head often retained; D1 and D2 bluish brown with whitish spots; caudal fin brown to dark grey; anal fin whitish
with inframarginal dark stripe to completely blackened; pectoral fin dusky with a pigmented dorsal margin; pelvic fin whitish to dusky
Diagnosis (Murdy, 1989;
Takita et al., 1999;
Polgar & Crosa, 2009):
total elements of D2 24-26;
total elements of anal fin 24-26; longitudinal scale count 61-79; predorsal scales 25-35; caudal fin length 17.9-23.3%SL; head length 25.0-30.4%SL;
length of D2 base 40.2-46.4%SL; 1st D2 element usually unsegmented and unbranched; lower jaw teeth notched (bifid).
Large blotches paler than background along the margin of the D1 membrane, behind the first 2-3 spines; pectoral fins with a dorsal blackened margin.
Species-specific sexual dimorphism: D1 spines more elongated in adult females (Polgar & Crosa, 2009; Swennen et al., 1995).
The genus is characterised by the greatly thickened epidermis of the head and dorsum, covered by dermal papillae; and by a rectangular piece of cartilage spanning the width of the
pelvic girdle (see drawing)
Diet:
a herbivorous benthic feeder: it scrapes the mud surface with side-to-side movements of the head (benthic diatoms, green algae: Khoo, 1966,
cited in Clayton, 1993; see the video clip). Where this species is syntopic with the congeneric
B. pectinirostris (e.g. in South-East Asia: Polgar & Crosa, 2009), the available studies did not take into account any possible difference between these two often confounded species (B. dussumieri sensu Takita et al., 1999 and Polgar & Khaironizam, 2008 = B. pectinirostris sensu Polgar & Crosa, 2009). The same is true in western India, where B. boddarti is sympatric with B. dussumieri (Murdy, 1989). However, all of these species exhibit almost identical feeding behaviours and probably have similar diets
Reproduction:
males jump to attract females inside the burrows to spawn (pers. obs.: Peninsular Malaysia); the details of its life
cycle have not been described in literature, but are probably similar to those of congeneric species
(see Reproductive behaviour and
B. pectinirostris)
|
|
Ecological notes:
locally very abundant along the marine fringe and in the pneumatophore zone of mangrove forests (Polgar & Crosa, 2009);
the adults dig burrows on the muddy banks of creeks, of river mouths, and on higher mudflats, usually not too far from the first row of trees
(Takita et al., 1999).
During low tide, B. boddarti shows ecological partition between larger individuals, found on unvegetated tidal mud and near the water's edge, and smaller individuals, on the forest floor and in the pneumatophore zone; a similar pattern was observed in different habitats of the same ecosystem in Periophthalmus gracilis and Oxuderces dentatus, while an apparently inverse pattern was observed in P. variabilis (Polgar & Bartolino, 2010). This pattern was observed in Boleophthalmus poti (= Boleophthalmus sp.; Polgar et al., 2010; Polgar et al., 2013) in Papua New Guinea, and in B. birdsongi in Australia (Takita et al., 2011). Ravi & Rajagopal (2007) studied its life history
left: Kuala Selangor, Peninsular Malaysia: forest marine fringe, mud banks of a creek; typical habitat of adult
B. boddarti (photo: G. Polgar, 1996)
right: Tg. Piai, Peninsular Malaysia: upper tidal flat with some trees of Rhizophora mucronata; here this species is very
abundant (photo: G. Polgar, 2006)
|
|
|
|
|
Distribution:
Indo-West-Pacific region, from the west coast of India (Mumbai, Maharashtra), eastwards to Sabah (northern Borneo) and Southern Viet Nam; type locality: Indian Ocean
(Murdy, 1989;
Bucholtz & Meilvang, 2005)
|
Remarks:
B. boddarti is extensively farmed and consumed in Thailand (see above)
(Clayton, 1993).
Chew & Ip (1992), Chew et al. (1998) and Ip et al. (2004) found a considerable resistance of this species to
environmental cyanide (produced by several organisms, such as cyanobacteria, fungi, plants, arthropods and algae: Ghosh et al., 2006) and sulfides (abundant in anoxic sediments). Both these toxic compounds inhibit aerobic cellular respiration via inhibition of the cytochrome oxydase. As a first rapid adaptive response, B. boddarti decreases the permeability of tissues to NH3 and
increase the production of NH3, thus increasing the pH of body fluids and converting toxic acids in ionized forms, which have a lower cell membrane permeability (Chew et al., 1998). Subsequently, specific enzymatic responses in the liver and muscle mediate detoxification through production of non-toxic thiocyanates, sulfates,
thiosulfates and sulfites in normoxic conditions. In hypoxic conditions, B. boddarti adopts O2-independent mechanisms, detoxifying sulfides to sulfane sulfur compounds and at the same time suppressing sulfide oxydase activity, thus freeing the limited supply of O2 (also due to the relatively high O2 affinity of mudskipper haemoglobin) for aerobic energy metabolism (Ip et al., 2004).
These findings suggest evolutionary adaptation to the anti-herbivory chemical
defenses adopted by the microalgae and cyanobacteria which this fish feeds on, and to the high environmental levels of these toxic chemicals
in the anoxic muds where these fishes dig burrows.
Interestingly, only specimens of B. boddarti >8 g body weight (about 80 mm SL: Khaironizam & Norma-Rashid, 2002) were able to tolerate sulfide exposure (Ip et al., 2004). This suggests that young (SL<80 mm) are not able to make use of hypoxic burrows during high tide, which is consistent with their presence of young at upper intertidal topographical levels relative to adults (Polgar & Crosa, 2009; Polgar & Bartolino, 2010), where substrate is submerged for a shorter time.
|
Photographs of Boleophthalmus boddarti:
|
A: a female of B. boddarti: note the elongated spines of D1 (photo: G. Polgar, Kuala Lumpur, Malaysia,
2006); B: a young specimen with the yellow D1 (photo: G. Polgar, Kuala Lumpur, Malaysia, 2006); C:
a close-up of a female (photo: G. Polgar, Kukup jetty, Peninsular Malaysia,
2006); D: another close-up of a female (photo: G. Polgar, Kukup jetty, Peninsular Malaysia,
2006); E: an agonistic encounter along the border of a territory (photo: G. Polgar, Kukup jetty, Peninsular Malaysia,
2006); F: a specimen "sieving" in a small pool the mixture of mud and algae scraped from the mud surface: note the ripples on the water surface made by the rapid movements of the mandible (photo: G. Polgar, Kukup jetty, Peninsular Malaysia,
2006); G: another close-up (photo: Akinori Kamiya "Yamaneko", Can Gio, Viet Nam,
2004;
© umisuzume
2006)*; H: B. boddarti preyed by the colubrid snake Cerberus rynchops
(photo: K. Khor, Kuala Gula, Malaysia, 2005); I:
a close-up of two fighting males (photo: K. Khor, Kuala Gula, Malaysia,
2005) - * with permission
|
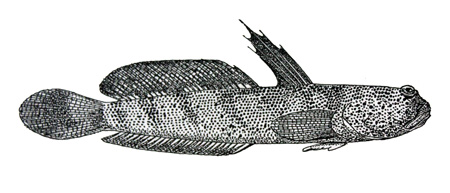
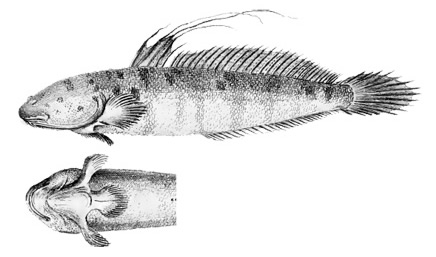
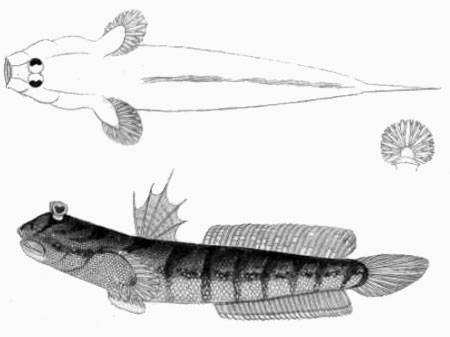
Drawings of Boleophthalmus boddarti:
A: Boleophthalmus boddarti (Pallas) (Koumans, 1953); B: Gobius striatus Bloch & Schneider (Bloch & Schneider, 1801);
C: B. boddaerti (Pallas) (Bleeker, 1983)*; D: Gobius boddarti Pallas (modified from Pallas, 1770); E: Boleophthalmus boddaerti
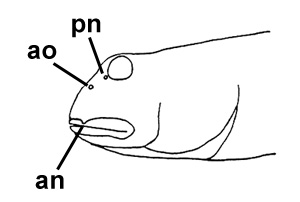
(Pallas) (Day, 1876; note that Apocryptes punctatus Day, 1867 does not include illustrations, but the description corresponds to B. boddarti); F: Gobius plinianus Hamilton (Hamilton, 1822); G: cephalic sensory and nasal pores of Boleophthalmus spp.: an = anterior nostril; ao = anterior oculoscapular canal pore;
pn = posterior nostril (modified from Murdy, 1989)*;
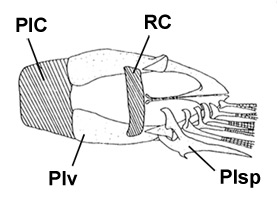
H: ventral view of the pelvis of B. boddarti (pelvic fin elements removed from left side: modified from
Murdy, 1989): PIC = pelvic intercleithral
cartilage; Plv = pelvis; RC = rectangular cartilage; Plsp = pelvic fin spine* - * with permission
|
Videos of Boleophthalmus boddarti:

click the image to download (right click --> Save as) or play
(left click) the videoclip
A feeding adult B. boddarti: the fish scrapes the biofilm made by benthic diatoms by pressing its mandible on the mud surface and moving the head from side to side. Scraping is facilitated by the rows of bifid teeth on the mandible, which are nearly horizontal (clip by Yuko Ikebe, Samut Prakarn, around the mouth of the Chao Phraya river, central Thailand, 2001)
|