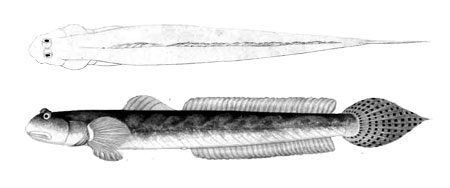
Common names:
Bangladesh |
Bengali |
chewa |
China |
Chinese Mandarin |
长身拟平牙虾虎鱼, 長身擬平牙鰕虎魚 (chang shen ni ping ya xia hu yu - long-bodied false flat-teethed goby), 矛状拟平牙虾虎鱼, 矛狀擬平牙鰕虎魚 (mao zhuang ni ping ya xia hu yu - lanceolate false flat-teethed goby) |
Denmark |
Danish |
keo-fisk |
Finland |
Finnish |
puikkokurkkija |
Germany |
German |
Lanzettgrundel |
Indonesia |
Bahasa Malaysia |
ikan njanjan |
Italy |
Italian |
saltafango allungato* |
Malaysia |
Bahasa Malaysia |
ikan senyenyen, belacak |
Taiwan |
Chinese |
尖尾鲨, 尖尾鯊 (jian wei sha - sharp-tailed shark) |
Viet Nam |
Vietnamese |
cá bống kèo |
United Kingdom |
English |
lanceolate goby |
* proposed name
|
|
|
Synonyms:
Eleotris lanceolata |
Bloch & Schneider, 1801 |
(replaced) |
Pseudapocryptes lanceolatus |
(Bloch & Schneider, 1801) |
(replaced, new combination) |
Apocryptes lanceolatus |
(Bloch & Schneider, 1801) |
(replaced, new combination) |
Gobius elongatus |
Cuvier, 1816 |
(replacement name*) |
Pseudapocryptes elongatus |
(Cuvier, 1816) |
(senior syn., new combination) |
Gobius changua |
Hamilton, 1822 |
(junior synonym) |
Apocryptes changua |
(Hamilton, 1822) |
(junior synonym) |
Apocryptes dentatus |
Valenciennes, 1837** |
(junior synonym) |
Boleophthalmus taylori |
Fowler, 1934 |
(junior synonym) |
Apocryptodon edwardi |
Fowler, 1937 |
(replacement name+) |
*substitute for Eleotris lanceolata
**In: Cuvier & Valenciennes, 1837
+substitute for Boleophthalmus taylori, sensu Fowler, 1937
Etymology:
'Pseudapocryptes' comes from the Greek 'pseudo' (false), and 'Apocryptes' which refers to the similarity to the
other oxudercine genus
'elongatus' in Latin means elongated, which refers to the eely shape of this fish
|
Maximum recorded length:
187 mm SL (Murdy, 1989)
24 cm TL (Dinh et al. 2007)
Live colouration (Murdy, 1989; pers. obs.: Peninsular Malaysia, southern Viet Nam):
colouration variable: ground colour dorsally yellowish to pale brown to reddish brown,
ventrally white, with no pigmentation on venter around the anal fin in smaller individuals; 6-8
dark brown diagonal saddle-like bars are frequently visible on flanks and dorsum; tiny
brown speckles may be present on dorsum; D1 translucent; D2 transparent with tiny brown
spots on rays and brown blotches basally and posteriorly in some individuals; caudal fin
yellowish to brownish with many brown speckles that can coalesce in wavy, broken lines;
anal, pectoral and pelvic fins translucent to yellowish to orange
Colouration on preservation (Murdy, 1989; pers. obs.: Bangladesh, Peninsular Malaysia, Viet Nam):
ground colour pale brown dorsally, yellow brown ventrally; dusky spots, bars and blotches may
be retained in recently preserved specimens; large dark brown blotches and bars on caudal peduncle and dark brown blotches on head frequently
retained, especially in smaller specimens (<100 mm SL); dorsal fins sometimes retain some dusky spots; all other fins, except caudal, mostly
transparent; patterns on caudal fin frequently retained
Diagnosis (Murdy, 1989):
total anal fin elements 28-31; body depth 9.9-13.6%SL; head length 14.8-22.0%SL; head depth 7.8-11.4%SL; pectoral fin length 10.4-13.4%SL;
pelvic fin length 9.0-11.5%SL; according to Murdy (1989) only
few brown spots are present on the body.
The genus is characterised by a longitudinal scale count of more than 150 scales
Diet:
benthic omnivorous feeders, prevalently feeding on phytoplankton (epibenthic diatoms, cyanobacteria); it
also feeds on small invertebrates, such as juvenile shrimps, and can live in aquarium for months feeding on Tubifex sp. worms. It apparently can perform three different feeding methods: (a) biting sediment and filtering food particles through the gill rakers; (b) pump-suspension-feeding (with an opercular beating rate 2-3 times faster than the normal breathing rate; Swennen et al., 1995); and (c) out of water on in extremely shallow water (partial emersion), mud scraping and filtering, with a side-to-side head movement similar to the behaviours of Boleophthalmus spp. and Z. confluentus; methods (a) and (b) may actually be two phases of the same behaviour: sediment collection and subsequent filtering through a flux of water (pers. obs.) (Swennen
et al., 1995; Bucholtz & Meilvang, 2005; Bucholtz et al., 2009)
Reproduction and life history:
the life history of this species is slightly different from the general model
proposed for all oxudercine gobies. After a planktonic stage along coast, larvae enter the intertidal zone from the sea
(recruitment), metamorphosing into juveniles and invading creeks, inlets and pools, where they grow for the next 8-9 months.
During the onset of the rainy season (May-June in southern Vietnam) adults migrate catadromously to the sea through the
creeks network (first males, then females), undergo sexual maturation, and eventually reach the breeding grounds,
which are probably located in mangrove areas and open mudflats of the lower intertidal and subtidal zones (Bucholtz et al., 2009).
In southern Vietnam (Dinh et al., 2007),
two spawning peaks occurred in July and October (which corrisponded to two recruitment peaks, 5 months apart),
during the rainy season (southern monsoon: April-November). Similarly, in India (Gangetic Delta), Hora
(1936) reported that the breeding
season is just before the southwest monsoon (July-November). Dinh et al. (2007) estimated a length at first maturity of 15-16 cm for both sexes (about 1 yr old), and a maximum longevity of about 4 years
|
|
Ecological notes (pers. obs.: Peninsular Malaysia, southern Viet Nam):
adults locally abundant on lower mudflats and in permanent creeks; young and subadults are abundant in mangrove forests, in aquatic microhabitats in the network of ephemeral inlets and tide pools, up to the supratidal zone (Bucholtz & Meilvang,
2005; pers. obs.); this species can aestivate inside vertical burrows (up to about 60 cm) during the dry season, when tide pools dry up
(Swennen et al., 1995; Hora, 1935)
left: Kuala Selangor, Peninsular Malaysia: an artificial bund channel
separates the mangrove forest from plantations or, as in this case, from secondary forest; here young
and subadults of Ps. elongatus are abundant
right: Kukup town: an artificial dike connected to a small mangrove forest: even here juveniles and adults are abundant
(photo: G. Polgar, 1996)
|
|
|
|
|
Distribution:
from the east coasts of India to South East Asia; its presence is not confirmed in
China and one single record is available from Kyushu Is., Japan (see drawing below; Tomiyama & Abe, 1958); type locality: Tranquebar, India (Murdy, 1989)
|
Remarks:
the discrimination between Ps. elongatus and the congeneric Ps. borneensis is mainly based on colouration, but several colouration
patterns with intermediate states can be found in nature (pers. obs.).
Murdy (1989) revised this species as Pseudapocryptes lanceolatus
(Bloch & Schneider, 1801): he designated a neotype (USNM 279321), since the type material is lost.
The name Pseudapocryptes elongatus
(Cuvier, 1816) is used after
Ferraris (1995), who showed that the name
Eleotris lanceolata Bloch & Schneider, 1801 (formerly junior synonym of Pseudapocryptes lanceolatus in Murdy, 1989)
was actually invalidated by Cuvier (1816: ICZN, 3rd ed., art. 59b), who replaced it with Gobius elongatus. Therefore, USNM 279321 is now the
neotype of Gobius elongatus Cuvier, 1816 (ICZN, 3rd ed., art 72e).
Ps. elongatus is extensively farmed in India and Southern Viet Nam, where it is largely consumed
(Bucholtz & Meilvang, 2005;
fishbase)
|
Photographs of Pseudapocryptes elongatus:
|
A: Ps. elongatus at low tide in a shallow pool on the higher mudflat (photo: G. Polgar, Kukup Jetty,
Peninsular Malaysia, 2007); B-D: close-ups of P. elongatus in aquarium (photos: G. Polgar, Kuala Lumpur, 2006); E: dissection: uncoiled gut of Ps. elongatus (photo in Bucholtz & Meilvang, 2005)*;
F: juvenile of Ps. elongatus in aquarium (photo: G. Polgar, Kuala Lumpur, 2006);
G: Ps. elongatus sold at the fish market of Can Tho, Viet Nam
(photo in Bucholtz & Meilvang, 2005)*; H: Ps. elongatus
in aquarium in a restaurant (photo: G. Polgar, Can Tho,
Viet Nam, 2006); I: Ps. elongatus roasted with veggies (photo: G. Polgar, Can Tho,
Viet Nam, 2006); J: a juvenile found inside a house artificial dike (photo: G. Polgar, Kukup town,
Peninsular Malaysia, 2007); K: vietnamese trap (Mekong Delta): the fisherman blocks one of the two openings of the fish burrow with a foot, and the other one with the trap, which has a small hole on the bottom (detail): the fish tries to escape and enters the trap (photo: Bucholtz & Meilvang, 2005)* - * with permission
|
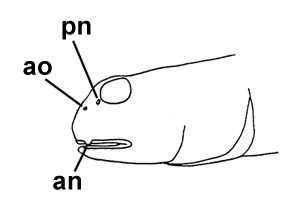
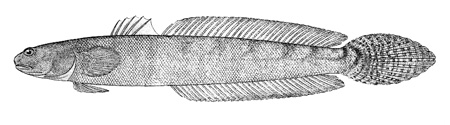
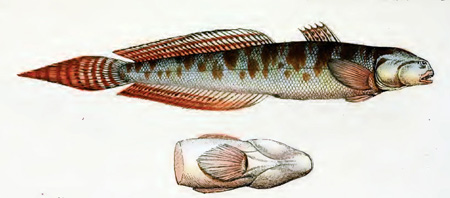
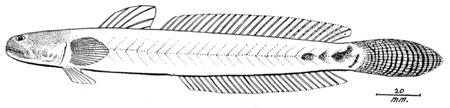
Drawings of Pseudapocryptes elongatus:
A: cephalic sensory and nasal pores of Pseudapocryptes spp.: an = anterior nostril; ao = anterior oculoscapular canal pore; pn = posterior nostril (modified from Murdy, 1989)*; B: Pseudapocryptes lanceolatus (Bloch & Schneider) (Koumans, 1953);
C: Pseudapocryptes lanceolatus (Bloch & Schneider) (Bleeker, 1983)*; D: Pseudapocryptes lanceolatus (Bloch & Schneider) (Tomiyama & Abe, 1958); Eleotris lanceolata Bloch & Schneider (Bloch & Schneider, 1801); F: Apocryptodon edwardi Fowler (Fowler, 1934; 1937); G: Apocryptes lanceolatus
(Bloch & Schneider) (Day, 1876); H:
Gobius changua Hamilton (Hamilton, 1822)- * with permission
|