Common names:
China |
Chinese Mandarin |
杜氏大弹涂鱼 () |
India, Gujarat |
Gujarati |
લેવતી (Levti) |
Iran, Hormozgan |
Farsi (Persian) |
eshlambo, neeshlambo, gel khorok |
Iraq, Oman |
Arabic |
kelb-el-daw, abou-shlembo, abu-shelamboo |
Italy |
Italian |
boleoftalmo di Dussumier* |
Pakistan |
Urdu |
gullo |
* proposed name
|
|
|
Synonyms:
Boleophthalmus dussumieri |
Valenciénnes, 1837* |
(senior synonym, original combination) |
Boleophthalmus dussumierei |
Valenciénnes, 1837* |
(emendatio incorrecta) |
Boleophthalmus dentatus |
Valenciénnes, 1837* |
(junior synonym) |
Boleophthalmus chamiri |
Holly, 1929 |
(junior synonym) |
Pseudapocryptes dentatus |
various authors |
(invalid combination) |
*In: Cuvier & Valenciénnes, 1837
Etymology:
'Boleophthalmus' depicts the capability of these fishes to rapidly raise their eyes above the level of their orbits, as if they were ejected out of their orbits: from 'bolê' (ejected), and 'ophthalmôn' (eye) (Cuvier & Valenciénnes, 1837)
the species is named after the French voyager and merchant Jean Jacques Dussumier, who collected the type material (Murdy, 1989)
|
Maximum recorded length:
187 mm TL (Randall, 1995)
289 mm TL (the largest recorded oxudercine goby: Abdoli et al., 2009)
Live colouration (Murdy, 1989;
pers. obs.: Iran, Hormozgan):
background colour bluish-gray to light brown on dorsum and flanks, with numerous dark brown speckles on the dorsal side of the head, gill covers, cheeks, muscular portions of pectoral fins, and on trunk; body ventral colouration paler;
darker saddle-like irregular vertical bars on dorsum, head and nape can be visible, but never below lateral midline;
D1 background light gray with numerous dark spots on the interradial membrane;
D2 background gray with columns of pale blue spots on the interradial membrane (basally larger), and few black spots anteriorly and basally; a narrow dark margin may be present, edged with orange;
caudal fin with dark gray to blackish rays and pale blue interradial membrane;
anal fin hyaline; pectoral fins pale brown to bluish with series of darker brown spots along rays; pelvic fins light brown, proximally darker
Colouration on preservation (Murdy, 1989;
pers. obs.: Iran, Iraq, Kuwait):
body background colour light brown to greyish blue or purplish; darker speckles on body and fins may be visible; saddle-like bars on dorsum not present; D1 and D2 background dusky to dark brown; D2 with whitish spots and dark margin often visible; caudal fin dusky to dark brown; anal fin and pelvic fins hyaline; pectoral fins light brown to hyaline
Diagnosis (Murdy, 1989):
total elements of D2 24-28;
caudal fin length 21.9-25.5%SL;
total elements of anal fin 24-27;
longitudinal scale count 103-185;
predorsal scales 48-56;
head length 23.6-28.7%SL;
length of D2 base 42.2-44.7%SL; first element of D2 usually unsegmented and unbranched; lower jaw teeth notched. The three upper canine teeth on both sides
of the symphysis are elongated, and obviously protrude in some preserved specimens; sexual dimorphism:
males have a higher D1 and larger canine teeth (Berg, 1949, cited by
Brian Coad; visited on 10/2009).
The genus is characterised by the greatly thickened epidermis of the head and dorsum, covered by dermal papillae; and by a rectangular piece of
cartilage spanning the width of the pelvic girdle (see drawing)
Diet:
Pankow & Huq (1979) and Sarker et al. (1980) described the gut content of Pseudapocryptes
dentatus from Shatt al-Arab, Iraq; Salih & Al-Jaffery (1980)
studied some morphological characters related to the feeding habits of B. dentatus (Iraq). This species principally feeds on epibenthic
diatoms, but also on cyanobacteria, filamentous algae, insects, crustaceans, nematodes and teleost fish eggs. The grazing behaviour, with typical
side-to-side head movements, is very similar to other congeneric species (have a look at a video clip of B. boddarti).
Reproduction:
Hoda & Akhtar (1985) and Hoda
(1986) studied the maturation and fecundity of B. dentatus
and B. dussumieri (Pakistan, Karachi coast) considering them as separate sympatric species. Courtship behaviour was described by Clayton
& Vaughan (1988): males perfom leaping behaviour, not unlike other congeneric species (pers. obs.). In Karachi (Pakistan), juveniles and young-of-the-year are found from September to December;
maturation starts at 60-65 mm SL in males and 70-75 mm SL in females, and spawning occurs twice per year during April-May and
July-September, or April-May and July-August (Hoda & Akhtar, 1985;
Hoda, 1986;
Hoda, 1987). In Kuwait, Clayton & Vaughan
(1988) reported the breeding season from March-April to August, and
the first appearance of larvae in mid-July.
|
|
Ecological notes:
This species can reach high densities on open tidal mudflats and on the mud banks of tidally influenced rivers, and is usually found at intermediate intertidal
levels (Polgar et al., 2009;
Clayton & Vaughan, 1988;
Clayton, 1985). B. dussumieri
builds peculiar systems of burrows and mud-walled polygonal territories (see photos C-E) [Clayton & Vaughan, 1982; 1986 (B. boddaerti = B. dussumieri,
misidentified); Clayton & Wright, 1989], which may have either the function
to reduce intraspecific aggressive interactions, or to facilitate the growth of benthic diatoms (sort of "farming ponds"), or both
[Clayton, 1987; this latter hypothesis was supported by Chen et al. (2009) for similar structures made by
B. pectinirostris in China]. The ethogram was described by Clayton
and Vaughan (1988), who also described a spatial partition between larger
territorial individuals and smaller not territorial or "errant" individuals, which exhibit gregarious behaviours. The thermal and seasonal behaviour
was described by Tytler and Vaughan (1983). These fishes are predated by numerous
predators, from wading birds to snakes and crocodiles (Clayton, 1988;
1993)
middle: Qeshm Island, Hormozgan, Iran: a densely populated tidal mudflat (photo: E. Kamrani, 2007, with permission)
|
|
|
|
Distribution:
Persian Gulf, Oman, Pakistan and west India; type locality of B. dentatus: Bombay (= Mumbai), Maharashtra, west India; type locality of B. chamiri: Strait of Hormoz, Hormozgan, Iran (Murdy, 1989)
|
Remarks:
Valenciénnes (in Cuvier & Valenciènnes, 1837)
considered B. dussumieri and B. dentatus as different species.
Day (1876) discriminated B. dussumieri from B. dentatus (sympatric populations from Mumbai, India; and Sind province plus Karachi, Pakistan) by the presence of shorter pelvic fins, a lower D1, and the presence of an interdorsal gap.
According to Berg (1949), the various nominal species were based on ontogenetic and sexual differences: B. chamiri was based on juveniles, which have fewer teeth;
B. dussumieri was based on females (smaller dorsal fins); and B. dentatus on males (cited by
Brian Coad; visited on 10/2009).
Hoda (1987) considered B. dussumieri and B. dentatus as
separate species, and even described their different length-weight relationships and the relative growth of several body parts; B. dentatus was also studied by Mansuri et al. (1982) in Gujarat, and by Soni and George
(1986). Subsequently, Murdy
(1989) synonymised all these species.
Several authors apparently misidentified B. dussumieri from the Persian Gulf as B. boddarti,
which is here absent (e.g. Al-Kadhomiy & Hughes,
1988; Clayton & Vaughan, 1988), but present along the west coasts of India
(e.g. Mumbai), where these two species may still be sympatric (Murdy, 1989).
Mirza et al. (1996) again discriminated B. dentatus from B. dussumieri
on the basis of D1 height, body depth, number of teeth and the presence/absence of a connection between the dorsal fins. More recently, Abdoli et al. (2009) found some unusually large specimens in southern Iran (see photo A, B), which closely resemble the B. dentatus described by Valenciénnes (see plate C); I examined similar large specimens in the fish collection of the Natural History Museum of London.
Polgar & Crosa (2009) proposed that the Malayan species described by Takita et al. (1999) and Polgar & Khaironizam (2008) in the Straits of Malacca as B. dussumieri, is in fact B. pectinirostris.
|
Photographs of Boleophthalmus dussumieri:
|
A: a very large specimen collected in Iran, Hormozgan, west of Bandar Abbas (the bar is 10 mm long; photo: G. Polgar, Hormozgan University, Marine Biology laboratory, 2007); B: a close-up of the head of the specimen in A: note the large hair-like dermal papillae (photo: G. Polgar, 2007); C:
polygonal territories on an tidal mudflat; researchers are collecting mudskippers by stick-beating, a traditional technique (photo: G. Polgar, Bandar Khamir, Hormozgan, Iran, 2007); D: another photo of the polygonal territories of B. dussumieri (photo: E. Kamrani, Qeshm Island, Hormozgan, Iran, 2007)*; E: another photo of polygonal territories (photo: D.A. Clayton, Sulaibikhat Bay, Kuwait, 1987); F-K: some shots from a sequence of an aggressive encounter: the two fishes are either in parallel, or reverse-parallel, or T formations, with both dorsal fins and caudal fin erected (Clayton & Vaughan, 1988); G: a "promeade" in parallel formation, with gaping mouths (a P. waltoni is also visible in the background); H: a violent bite in a T formation; I-K: "tail-beating", a behaviour that was not reported by the cited authors; L: a close-up in
aquarium (photo: G. Polgar, Hormozgan University, Marine Biology laboratory, 2007) - * with permission
|
Drawings of Boleophthalmus dussumieri:
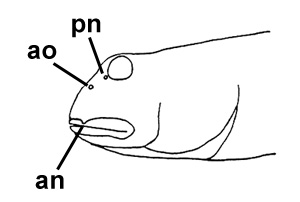
A: cephalic sensory and nasal pores of Boleophthalmus spp.: an = anterior nostril; ao = anterior oculoscapular canal pore;
pn = posterior nostril (modified from Murdy, 1989)*;
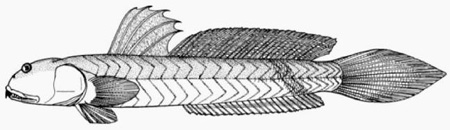
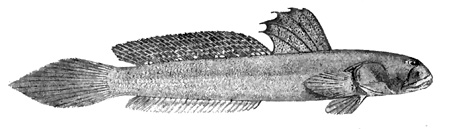
B: drawing of B. dussumieri by Susan Laurie-Bourque (Brian W. Coad's Personal Website)*; C: Boleophthalmus dussumieri Valenciénnes (above)
and B. dentatus Valenciénnes (below) (Cuvier &
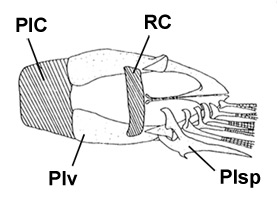
Valenciénnes, 1837); D: ventral view of the pelvis of B. boddarti (pelvic fin elements removed from left side:
modified from Murdy, 1989): PIC = pelvic
intercleithral cartilage; Plv = pelvis; RC = rectangular cartilage; Plsp = pelvic fin spine (modified
from Murdy, 1989)*; E:
Boleophthalmus dentatus Valenciénnes (Day, 1889) - * with permission
|