|
Periophthalmus sp. has a ‘ramp’ retina: the distance from the centre of the lens to the retina increases
in the vertical direction (Karsten, 1923; Munk, 1970); this contemporaneously allows it to focus on near objects in the dorsal portion of the retina, and distant objects
in the ventral one.
Higher density of photoreceptors are found in the dorsal part of the
retina.
In Boleophthalmus sp. and Periophthalmus sp. the retina is rich in cones and strongly regionalised:
photoreceptors are arranged in horizontal bands dorsally to the fundus (Munk, 1970; Kotrschal, 1999; Schwab, 2003).
This would
allow a higher resolution and a lower perceptive threshold for movement, especially on the horizontal plane and at right
angles to the anatomical axis of the eye, probably increasing the ability to detect small preys (Clayton, 1993).
A regionalised retina had also been found in other unrelated gobies and blennies, and seems to be an adaptation to benthic life in small fishes (Kotrschal, 1999).
In Periophthalmus sp. the optic nerve fibres do not converge on the optic disc as in other fishes, but are spread out over a larger part of the retinal surface, possibly to avoid reducing the sensitivity of the areae (Munk, 1970).
Some species of the genus Periophthalmus present high concentrations of pigments (Graham, 1971)
to protect the retina from the intense illumination of the exposed subaerial environment (e.g. on the mudflat).
Other species instead present a strongly reflective argentea fibrosa (Karsten, 1923), an adaptation to dark environments
(e.g. inside the forest, or at night).
The eyes of some species of Periophthalmus living in the high mangrove forest visibly
reflect the light of a torch in the darkness, even from within their burrows (Polgar, 2004).
Aerial vision could be important also for homing
behaviour, and some studies suggest that Periophthalmus sobrinus (= P. argentilineatus) is capable of using the sun to orient (Berti et al., 1992; 1994).
Also mechanoreception apparently has an important role.
For instance, P. chrysospilos seems to orient by means of a rheotactic
response at ebb tide, when it forms coordinated schools of individuals to follow the water edge (Polgar, 2004).
|
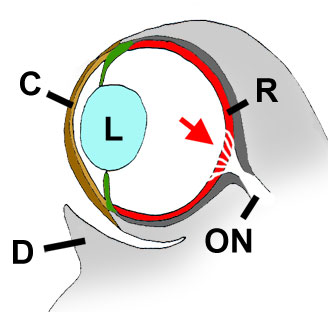
Cross section through a mudskipper's eye. C= cornea (including c. propria, or inner/proximal lamella;
and c. conjunctiva, or outer/distal lamella). L= lens; R= retina; ON= optical nerve; D= dermal cup. Red arrow= retinal area
where the optic nerve fibres are spread out.
Modified from Graham, 1971, with permission
|