|
Common names:
China, Taiwan |
Chinese Mandarin |
弹涂鱼, 彈塗魚 (tan tu yu - Mudskipper) |
Korea |
Korean |
말뚝망둥어 |
Japan |
Japanese |
トビハゼ (Tobihaze) |
Hong Kong |
Cantonese |
nai hau - Mud Monkey |
Hong Kong |
English |
Mudskipper |
Italia |
Italian |
perioftalmo modesto* |
Sweden |
Swedish |
Kantonesisk Slamkrypare |
Viet Nam |
Vietnamese |
Cá thoi loi |
* proposed name
|
|
|
Synonyms:
|
Cyprinus cantonensis |
Osbeck, 1765 |
(pre-linnean, invalid) |
|
Periophthalmus cantonensis |
(Osbeck, 1765) |
(pre-linnean, new combination, invalid) |
|
Periophthalmus modestus |
Cantor, 1842 |
(senior syn., original combination) |
Etymology:
'Periophthalmus' is a compound name from the Greek 'peri' (around), and 'ophthalmôn' (eye), which refers to the wide
visual field of these species
'modestus' means 'modest' in Latin, which probably refers to the lack of striking colouration
(Murdy, 1989)
|
Maximum recorded length:
80.2 mm SL (Lee et al., 1995)
Live colouration
(Murdy, 1989;
Lee et al., 1995;
Baeck et al., 2008; pers. obs.: from available photos):
background colour grey to light brown, with tiny whitish spots on cheeks and opercula, and several small black speckles on flanks; ventrally whitish; head
ventrally white; several dorsal dark brown saddle-like bars may be visible; D1 grey to brownish, darker inframarginally,
and a narrow whitish to transparent margin. D2 transparent with a grey to brownish medial stripe and dark speckles basally on rays; pectoral and caudal fins dusky,
with series of dark speckles along rays in some specimens; anal fin white; pelvic fins ventrally whitish, dorsally lightly pigmented in some specimens
Colouration on preservation
(Murdy, 1989; Lee et al., 1995; Baeck et al., 2008; pers. obs.: Taiwan, Japan):
ground colour grey to brownish on dorsum and flanks, ventrally whitish; white speckles on head, dark speckles on flanks, and dorsal banded pattern may be preserved; D1 background brownish,
with a pale to transparent margin;
D2 transparent with a dark medial stripe and dark speckles basally on rays; caudal and pectoral fins dusky with series of
dark speckles on rays; pelvic fins ventrally white, dorsally lightly pigmented; anal fin white
Diagnosis (Lee et al., 1995):
D1 X-XIV; total D2 elements 11-13; total anal fin elements 11-12; longitudinal scale count 75-84; predorsal scale count 35-40; length of D1 base 20.4-25.3%SL; pelvic fins only partially united by a basal membrane, and with a prominent frenum uniting the two spines; D1 with no spots; D2 with a dark medial stripe and basal series of dark speckles on rays; dorsal fins not connected by membrane; caudal fin broad and rounded; no elongated spines; small black speckles on flanks.
The genus is yet undefined by synapomorphies
Diet:
no published study is available
|
|
Reproduction:
in south Korea the breeding season is from early May to July (Baeck
et al., 2008), while in southern Japan (Ariake Bay) is from May to August (Ishimatsu et al., 2007). Reproductive males dig burrows on unvegetated mudflats. To attract females, they change
their body background colour from grey to pinkish, raise and wave their tails, and make vertical jumps with the spread fins. When the female approaches, the male
induces her to enter the
burrow by slow, wiggling movements (Matoba & Dotsu, 1977;
Baeck et al., 2008). Reproductive burrows have 2-3 narrow openings with no turrets, and a sub-vertical shaft which ends into an upturned tract, where the eggs
are laid (Kobayashi et al., 1971; Matoba & Dotsu, 1977; Ishimatsu et al., 2007).
The cohabitation prolongs only until spawning, then the female leaves the nest, and the male guards the eggs for about one week,
always remaining nearby the nest and maintaining an air phase inside the egg chamber; during high tide, when the nest is submerged, the male
remains inside the burrow. When the eggs are ready to hatch, the male floods the egg chamber to let the larvae leave the burrow during ebb tide
(Ishimatsu et al., 2007). The larval development was
described by Kobayashi et al., 1972: the larvae are
planktonic until 30-40 days after hatching, then they enter the intertidal zone again (recruitment), become benthic and rapidly metamorphose (settlement)
middle: a couple of P. modestus during courtship: male on the left
photo: Kyoto, southern Japan, 2003 (©
BlueNativeFactory, 2008, with permission)
|
|
|
|
Ecological notes:
very abundant on open mudflats and mouth of rivers, on level and unvegetated mud deposits. P. modestus is distributed in temperate regions, and it hybernates during winter, remaining inside burrows (in southern Japan, from November to March: Kobayashi et al., 1971). In south Korea, P. modestus is active from late April to October (Baeck et al., 2008).
During the hot season, burrows are apparently occupied and maintained only for the reproduction; for the rest, P. modestus opportunistically utilizes crevices and burrows of different animals as temporary shelters during low tide (Oshiro et al., 2005; Baeck et al., 2008). During ebb tide, P. modestus follow the water edge to feed on the mudflat, while during flood
tide they perch on emerged objects, keeping out of water; this behaviour is also temperature-dependent: if it is too cold, fishes remain inside burrows (Ikebe & Oishi, 1996). This species is frequently sympatric with Boleophthalmus pectinirostris, and P. magnuspinnatus, which usually lives at higher intertidal levels (Baeck et al., 2008).
middle: a tidal flat inhabited by a colony of P. modestus in Tamagawa, southern Japan (photo: A. Kamiya "Yamaneko", © umisuzume)
|
|
|
|
Distribution (Murdy, 1989; Wang et al., 2006):
from southern Japan to Korea, southward to Hong Kong;
type locality: Chusan, China
|
|
|
Remarks:
In China, P. modestus was confused with P. magnuspinnatus for a long time after the description of this latter species in Korea (Lee et al., 1995). A molecular study (Wang et al., 2006) verified the presence of both species in China
|
Photographs of Periophthalmus modestus:
|
|
A: close-up of P. modestus (photo: 'hakofugu', port of Onomichy, Giappone, 2003: fishingforum - link removed); B: a specimen in aquarium: notice the structure of the pelvic fins ('chinya', Kumamoto prefecture, Giappone, 2004: fishingforum);
C: another close-up: notice the erected D1; D, E:
individuals taking shelter on emerged trunks during high tide; F: P. modestus spitting mud during burrow excavation; G: a burrow with a large opening: notice the round mud gobbets produced during excavation; H: an individual (possibly a male guarding eggs) threatening a crab; I: another individual inside a narrow burrow (photos C-I: A. Kamiya "Yamaneko", Edogawa, Giappone, 2001, © umisuzume, 2007); J: a male (left) courting a female (right), trying to attract her inside the nest (photo: Ariake Sound, Kyoto, 2003 © BlueNativeFactory, 2008)*; K: an individual in aquarium threatening
another one in the background; L: another individual in aquarium (photos K, L: M. Tanabe, © Marli's mudskipperland, 2007) - * with permission
|
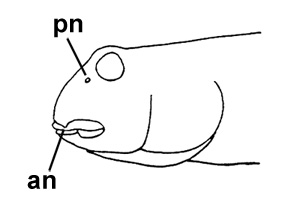
Drawings of Periophthalmus modestus:
left: cephalic sensory and nasal pores of Periophthalmus spp.: an= anterior nostril; pn= posterior nostril
(modified from Murdy, 1989)* - * with permission
|