|
Many oxudercine species face rapid and drastic changes
of environmental salinity and are euryhaline.
Osmoregulation in hyperosmotic environment (i.e. sea water) is realised
through the accumulation of free amino acids (FAA) and ammonia (NH3) in muscles (Iwata et al.,
1981). The increased osmolality of body fluids limits the passive intake
of ions. Salts in excess are excreted through the gills by means of the
"chloride cells" (Evans et al., 1999).
In Periophthalmus modestus, and probably in all congeneric species that have strongly reduced gills, this process takes
place also in some cutaneous areas (i.e. behind pectoral fins) through
specialized, mitochondria-rich cells (Yokota et al., 1997; Sakamoto
et al., 2000; Sakamoto & Ando, 2002).
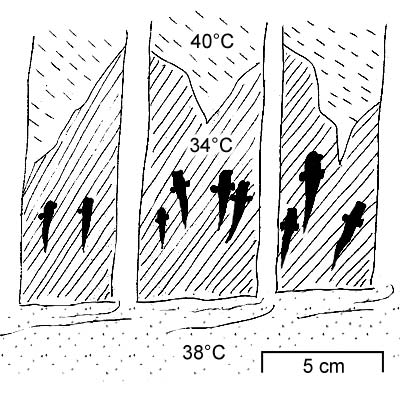
Osmoregulation at low salinity levels instead (hyposmotic environment)
seems at least partly behavioural: emergence would compensate for the excessive
hydration through cutaneous evapotranspiration (Clayton, 1993).
The resistance to dehydration upon emergence, related to hyperosmotic osmoregulative capabilities (Evans et al. 1999), was also measured in few amphibious species, which presented
an evaporative water permeability comparable to frogs (Gordon et al. 1969, 1978). The structure of skin and some particular mucous secretions can partly account for such
capabilities, performing a trade-off between cutaneous respiration and water losses; species with different degrees of adaptation to terrestriality present different skin histologies (Zhang et al. 2000; 2003).
|
|
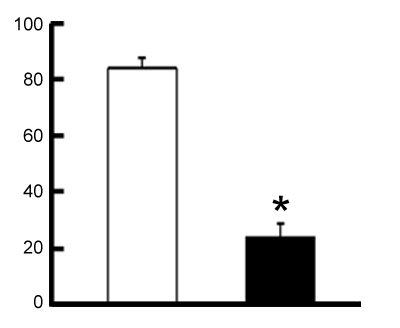
Percentages of chloride cells with Con-A binding area
(i.e. with open apycal crypts, actively secreting salts) in 4 fishes immersed in 10mmol*l^-1
CaCl2 solution (left), and in freshwater (right).
*Significance level P<0.001.
Drawing: G. Polgar, redrawn from Sakamoto & Ando, 2002, with permission from the first author
|