|
Periophthalmus sp. ha una retina ‘a rampa’: la distanza dal centro del cristallino e la retina aumenta lungo la direzione verticale (Karsten, 1923; Munk, 1970); ciò permette di mettere a fuoco contemporaneamente oggetti vicini sulla porzione dorsale della retina, ed oggetti distanti su quella ventrale.
Nella parte dorsale della retina si trova una maggiore densità di fotorecettori.
In Boleophthalmus sp. ed in Periophthalmus sp. la retina è ricca di coni e fortemente regionalizzata: dorsalmente al fundus i fotorecettori sono disposti in bande orizzontali (Munk, 1970; Kotrschal, 1999; Schwab, 2003).
Ciò permetterebbe una maggiore risoluzione ed una minore soglia di percezione per il movimento, specialmente sul piano orizzontale e ad angoli retti rispetto all’asse anatomico dell’occhio, probabilmente aumentando così l’abilità di individuare piccole prede (Clayton, 1993).
Una retina regionalizzata è stata anche trovata in altre specie di gobidi e di blennidi e sembra essere un adattamento alla vita bentica in piccoli pesci (Kotrschal, 1999).
In Periophthalmus sp. le fibre del nervo ottico non convergono sul disco ottico come nella maggioranza degli altri pesci, ma sono sparse su una porzione più ampia della superficie della retina, forse per evitare di ridurre la sensibilità delle areae (Munk, 1970).
Alcune specie del genere Periophthalmus presentano elevate concentrazioni di pigmento (Graham, 1971) per proteggere la retina dalle elevate intensità di illuminazione degli ambienti subaerei (per es. sulle piane tidali).
Altre specie invece presentano una argentea fibrosa a forte potere riflettente (Karsten, 1923), un adattamento ad ambienti scarsamente illuminati (per es. all’interno della foresta, oppure di notte).
Gli occhi di alcune specie di Periophthalmus che vivono in foresta alta riflettono visibilmente la luce di una torcia al buio, persino dall’interno dei loro cunicoli (Polgar, 2004).
La visione aerea potrebbe svolgere un ruolo importante anche nei comportamenti di orientamento e di homing, ed alcuni studi suggeriscono che Periophthalmus sobrinus (= P. argentilineatus) sia in grado di utilizzare il sole per orientarsi (Berti et al., 1992; 1994).
Un ruolo importante sembra venga ricoperto anche dalla percezione meccanosensoriale. P. chrysospilos ad esempio, sembra orientarsi per mezzo di una risposta reotattica alla marea calante, quando forma banchi coordinati di alcune decine di individui per seguire il bordo dell’acqua (Polgar, 2004). |
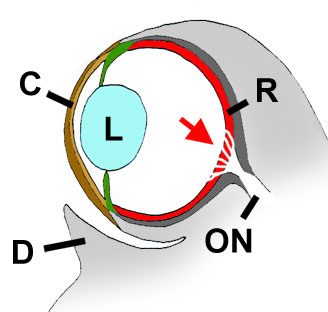
Sezione trasversale attraverso un occhio di saltafango. C= cornea (incluse la c. propria, o lamella interna/prossimale; e c. conjunctiva, o lamella esterna/distale). L= cristallino; R= retina; ON= nervo ottico; D= coppa dermica. Freccia rossa= area retinica dove sono sparse le fibre del nervo ottico. Modificato da Graham, 1971, con il permesso dell’autore
|